

1_2_Lektsia_Obmen_uglevodov_chast_2
.pdfБиохимия углеводов
Часть II. ОСНОВНЫЕ ПУТИ КАТАБОЛИЗМА ГЛЮКОЗЫ

Содержание
1.Гликолиз.
2.Пентозо-фосфатный путь.

Окисление глюкозы до СО2 и Н2О (аэробный распад). Аэробный распад глюкозы можно выразить суммарным уравнением:
С6Н12О6 + 6О2 → 6СО2 + 6Н2О + 2820 кДж/моль. Этот процесс включает несколько стадий:
•аэробный гликолиз (в узком смысле) – процесс окисления глюкозы с образованием двух молекул пировиноградной кислоты (ПВК);
•общий путь катаболизма, включающий окислительное декарбоксилирование ПВК в ацетил-КоА, и его дальнейшее окисление в ЦТК;
•окислительное фосфорилирование, сопряжѐнное с реакциями дегидрирования (в самом гликолизе и общем пути катаболизма), происходящими в процессе распада
глюкозы.
Анаэробный распад (анаэробный гликолиз)
В определѐнных ситуациях обеспечение кислородом тканей может не соответствовать их потребностям. Например, на начальных стадиях интенсивной мышечной работы
при стрессе сердечные сокращения могут не достигать нужной частоты, а потребности
мышц в кислороде для аэробного распада глюкозы велики. В подобных случаях включается процесс, который протекает без кислорода и заканчивается образованием молочной кислоты (лактата) из ПВК. Этот процесс называют анаэробным распадом, или анаэробным гликолизом. Анаэробный распад глюкозы энергетически
малоэффективен, но именно он может стать единственным источником энергии для
мышечной клетки в описанной ситуации. В дальнейшем, когда снабжение мышц кислородом будет достаточным в результате перехода сердца на ускоренный ритм, анаэробный распад переключается на аэробный.

Реакции аэробного гликолиза
Глюкозо-6-фосфат, образованный в результате фосфорилирования глюкозы с участием АТФ, в ходе следующей реакции превращается в фруктозо-6-фосфат. Эта обратимая реакция изомеризации протекает под действием фермента фосфоглюкоизомеразы.
Затем следует ещѐ одна реакция фосфорилирования. В ходе этой реакции, катализируемой фосфофруктокиназой, фруктозо-6-фосфат превращается в фруктозо- 1,6-дифосфат. Данная реакция, так же, как гексокиназная, практически необратима, и, кроме того, она наиболее медленная из всех реакций гликолиза. Реакция, катализируемая фосфофруктокиназой, определяет скорость всего гликолиза, поэтому, регулируя активность фосфофруктокиназы, можно изменять скорость катаболизма
глюкозы.
Фруктозо-1,6-дифосфат далее расщепляется на 2 триозо-фосфата: глицеральдегид- 3-фосфат и дигидроксиацетонфосфат (ДГАФ). Реакцию катализирует фермент фруктозодифосфатальдолаза, или просто альдолаза. Эта реакцию обратима. Продукты реакции альдольного расщепления – изомеры. В последующих
реакциях гликолиза используется только глицеральдегид-3-фосфат, поэтому ДГАФ
превращается с участием фермента триозофосфатизомеразы в глицероальдегид-3- фосфат.
Вместе с тем, при избытке глюкозы ДГАФ направляется на синтез липидов как источник их глицеринового скелета*.
* См. следующий раздел «Обмен липидов».
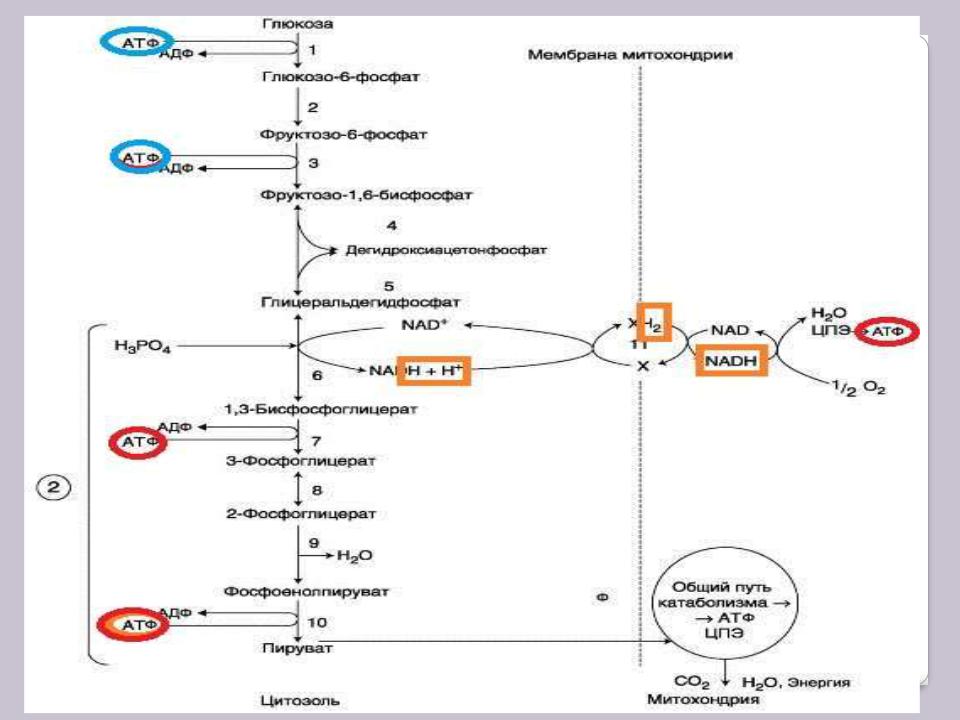

Окисление цитоплазматического NADH+Н+ в ЦПЭ. Челночные системы
NADH + Н+ (иногда для простоты пишут просто NADH2) oбразующийся при окислении глицеральдегид-3-фосфата в аэробном гликолизе, подвергается окислению путѐм переноса атомов водорода в ЦПЭ. Однако митохондриальная мембрана для него непроницаема. Перенос водорода через мембрану происходит с помощью специальных «челночных» механизмов: водород транспортируется через мембрану при участии пар субстратов, связанных соответствующими дегидрогеназами, т.е., с обеих сторон митохондриальной мембраны находится специфическая дегидрогеназа.
Известны 2 челночных механизма для переноса водорода. В первой из них водород от NADH в цитозоле передаѐтся на ДГАФ ферментом глицерол-3- фосфатдегидрогеназой (NAD-зависимой). Образованный в ходе этой реакции глицерол-3-фосфат (другое название – α-глицерофосфат), окисляется далее ферментом внутренней мембраны митохондрий – глицерол-3- фосфатдегидрогеназой (FAD-зависимой). Затем протоны и электроны с FADH2 переходят на убихинон, и далее по ЦПЭ.
А-глицерофосфатный челночный механизм работает в клетках белых мышц и гепатоцитов. Однако в клетках сердечных мышц митохондриальная глицерол-3-фосфат-дегидрогеназа отсутствует.
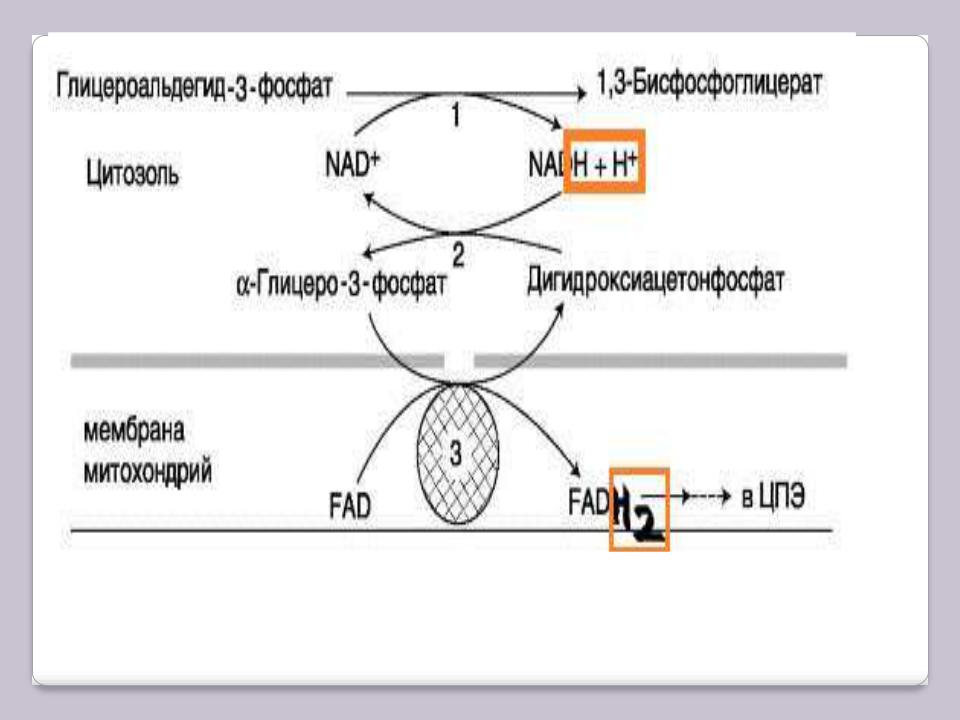
А-глицерофосфатный челночный механизм
1– глицеральдегид-3-фосфатдегидрогеназа;
2– глицерол-3-фосфатдегидрогеназа;
3– глицерол-3-фосфат-дегидрогеназа.

Малат-аспартатный челночный механизм, в
которой участвуют яблочная кислота (малат), цитозольная и митохондриальная малат-дегидрогеназы, является более универсальной.
В цитоплазме NADH восстанавливает щавелево-уксусную кислоту (оксалоацетат) в малат, который при участии переносчика проходит в митохондрии, где окисляется в оксалоацетат NAD-зависимой малатдегидрогеназой. Восстановленный в ходе этой реакции NAD отдаѐт водород в ЦПЭ. Однако образованный из малата оксалоацетат выйти самостоятельно из митохондрий в цитозоль не может, так как мембрана митохондрий для него непроницаема. Поэтому оксалоацетат путѐм трансаминирования* превращается в аспарагиновую кислоту (аспартат), который и транспортируется в цитозоль, где снова превращается в оксалоацетат.
Обе челночные системы отличаются по количеству синтезированного АТФ. В первой системе соотношение Р/O равно 2, так как водород вводится в ЦПЭ на уровне KoQ. Вторая система энергетически более эффективна, так как передаѐт водород в ЦПЭ через митохондриальный NAD+ и соотношение Р/О близко к 3.
* см. раздел «Азотистый обмен на II курсе»
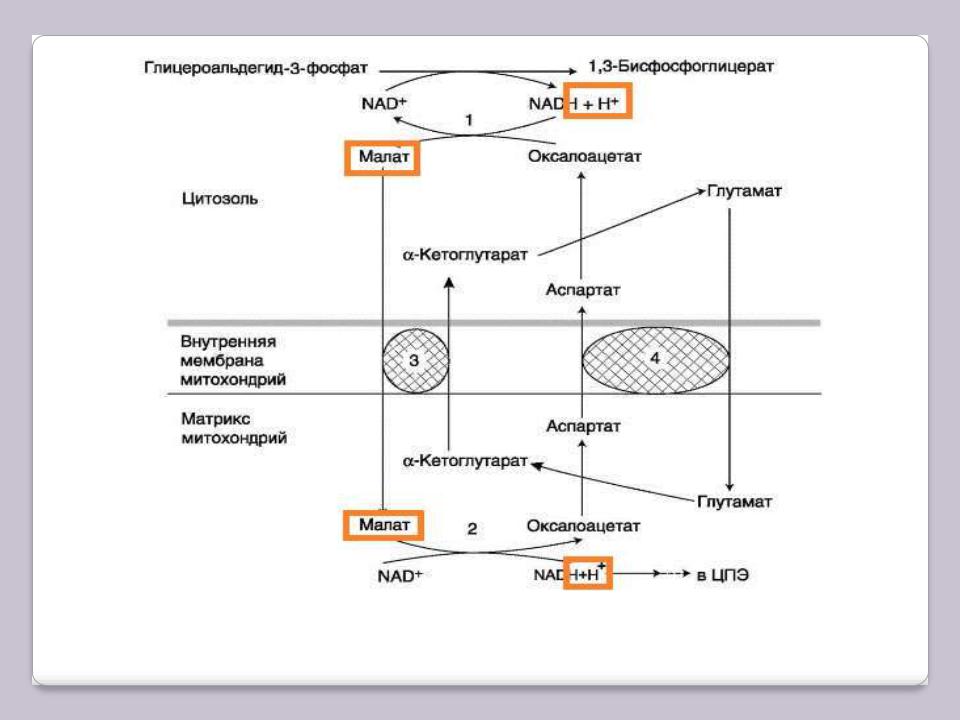
Малат-аспартатный челночный механизм
1, 2 – окислительно-восстановительны реакции, обеспечивающие транспорт водорода из цитозоля в митохондрии на ЦПЭ; 3, 4 – транслоказы, обеспечивающие транспорт α-кето-глутарата, аспартата и глутамата через мембрану митохондрий.
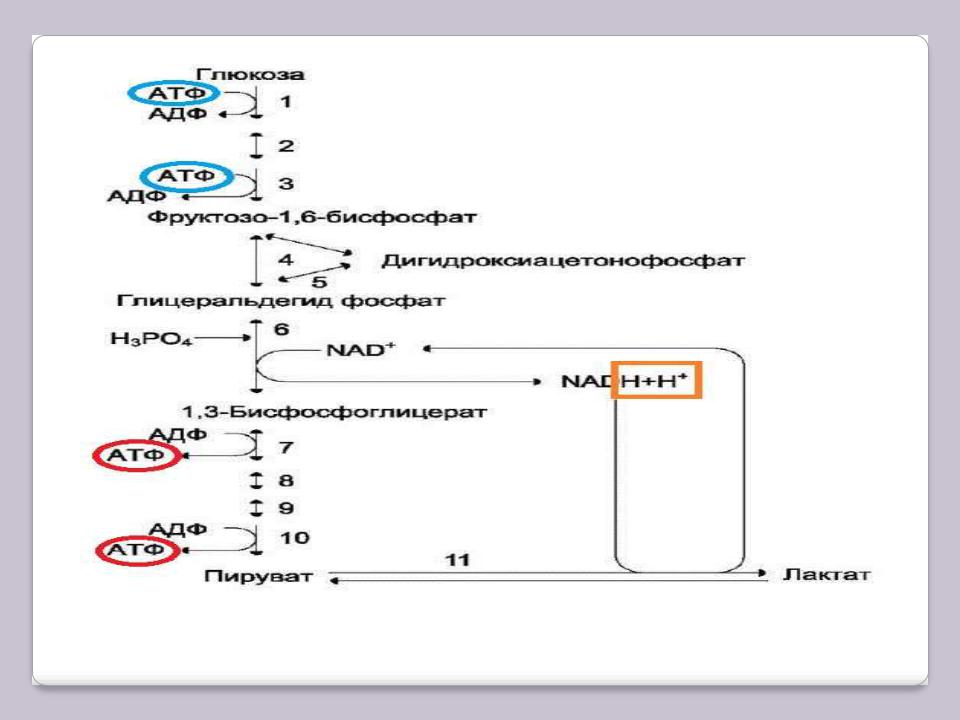
Анаэробный гликолиз
6, 11 – гликолитическая оксидоредукция.
Голубым цветом обведены затраты АТФ, красным – получение, оранжевым – водороды в составе восстановленных коферментов.