

Презентации / 2-02- Нарушения КОС
.pdfЭффективность и вклад основных буферных систем
вобеспечение стабильности рН
-эффективность зависит от:
•емкости буферной системы,
•скорости вступления в реакции нейтрализации сдвигов КОС;
•реактивности организма, проявляемой к отдельным компонентам буферной системы;
-вклад буферных систем в нейтрализацию сдвигов рН при добавлении к 1 литру плазмы 28 ммоль Н+ и смещении его с 7,4
до 7,0:
Бикарбонатная буферная система - 18 ммоль
Фосфатная буферная система - 0,3 ммоль
Гемоглобиновая буферная система - 8,0 ммоль
Белковая буферная система - 1,7 ммоль
(Беляевский А.Д., Монченко Г.Д., 1997)

Эффективность и вклад основных буферных систем в обеспечение стабильности рН
-кровь составляет лишь 20% общей буферной емкости организма;
-по резервной мощности и скорости гидрокарбонатная буферная система стоит на первом месте, обеспечивая 53% общей буферной емкости крови (35% - гидрокарбонат плазмы и 18% - внутриэритроцитарный гидрокарбонат);
-некарбонатные системы обеспечивают остальные 47%: фосфаты - 5%
белки - 7 - 10%
-емкость гемоглобинового буфера составляет 2/3 (75% Иванов И.И., 1969) всех химических буферов крови, однако в функциональном отношении активна лишь 1/3 (около 250 г), обеспечивая лишь 35% буферирования крови.
(Зильбер А.П., 1984; Жизневский Я.А., 1994; Hruska K., 1986)

РОЛЬ СИСТЕМЫ ВНЕШНЕГО ДЫХАНИЯ В РЕГУЛЯЦИИ РН
-удаляют СО2, образующийся в ходе метаболических процессов и нейтрализации кислых валентностей;
-смещают равновесие реакции Н+ + НСО3- = Н2СО3 = Н2О + СО2 вправо;
-на величину альвеолярной вентиляции влияют оба
компонента бикарбонатного буфера (СО2 - непосредственно, НСО3- - опосредованно);
-дыхательная адаптация к метаболическим нарушениям рН является быстрой, начинаясь через 1-2 минуты после сдвига КОС, достигает максимума ч-з 12-24 часа;
-компенсаторные механизмы вентиляции в обеспечении стабильности КОС временны, они редко восстанавливают рН до полностью нормальных значений;
-доля легких в восстановлении рН составляет 50-70%;
(Аганезова Е.С., 1980; Нruska K., 1987; Horne M.M. et al., 1999)

РОЛЬ СИСТЕМЫ ВНЕШНЕГО ДЫХАНИЯ В РЕГУЛЯЦИИ РН
-гипервентиляция при метаболическом ацидозе более эффективна, чем при метаболическом алкалозе;
-избыток Н+ вызывает развитие тахи- и гиперпноэ;
-ацидемия в 4-5 раз повышает объем альвеолярной вентиляции
(при снижении НСО3- на 1,0 ммоль/л напряжение рСО2 снижается на 1,1 мм рт.ст.);
-даже при выраженном метаболическом ацидозе стимуляция вентиляционной функции легких достигает предела при достижении раСО2 10-20 мм рт.ст.;
-в ответ на алкалемию снижение вентиляционной функции достигает 50-75% от исходного уровня, а объем дыхательной
компенсации метаболического алкалоза ограничен раСО2 = 55 мм рт.ст.;
-раСО2 = 65 мм рт.ст. - раО2 < 80 мм рт.ст. - является стимулятором дыхательного центра.
(Малышев В.Д., 2000; Тino G., Grippi M.A., 1997)

РОЛЬ ПОЧЕК В РЕГУЛЯЦИИ РН
- основной механизм поддержания КОС плазмы крови:
•реабсорбция профильтровавшегося НСО3- из первичной мочи;
•экскреция кислых эквивалентов нелетучих кислот;
-экскреция Н+ не осуществляется до тех пор, пока не реабсорбируется профильтровавшийся НСО3-;
-в проксимальном извитом канальце реабсорбируется 80-90% профильтровавшегося NaНСО3-, без секреции в этом отделе Н+;
-реабсорбция НСО3- эквимолярна секреции в этом отделе Н+, в связи с чем здесь не происходит истинной секреция протона;
-в прямой части проксимального почечного канальца и петле
Генле реабсорбируется 5-20% NaНСО3-, профильтровавшегося в первичную мочу.
(Forman J.W., 1986; Hruska K., 1987)

РОЛЬ ПОЧЕК В РЕГУЛЯЦИИ РН
-в дистальных отделах нефрона происходит полная
реабсорбция 1-5% NaНСО3- - лишь после этого возможно снижение рН мочи;
-в дистальном канальце и собирательных трубочках происходит итоговая секреция Н+:
•АТФ-зависимый Н+-насос
•плотные контакты эпителиоцитов, предотвращают выход Н+
винтерстиций
•отсутствие карбоангидразы на люминальной поверхности,
предотвращают дегидратацию Н2СО3 (концентрация Н+ увеличивается в 1000 раз по сравнению с плазменной);
-99,9% Н+ буферировано:
1/3 - фосфатами, |
2/3 - аммиаком; |
- Н+ могут буферироваться мочевой кислотой, сульфатами, кетокислотами, но их роль в этом процессе невелика.
(Forman J.W., 1986; Hruska K., 1987)

* Регуляция КОС почками (1) Реабсорбция бикарбонатов
просвет канальца
клубочек
NaHCO3
H2CO3
H2O CO2
|
|
кровеносный |
|
|
сосуд |
эпителий |
|
|
канальца |
|
|
|
|
NaHCO |
H2O + CO2 |
|
|
|
карбоан- |
|
|
гидраза |
|
|
H2CO |
Na } |
|
3 |
|
|
|
|
Н+ |
HCO3 |
HCO3- |
|
- |
+ |
|
|
|

РОЛЬ ПОЧЕК В РЕГУЛЯЦИИ РН
-для секреции в дистальных канальцах Н+ необходимо поступление в них «транспортного максимума» натрия, что необходимо для сохранения электронейтральности при секреции Н+;
-Н+ в дистальных канальцах транспортируется против концентрационного градиента;
-буферные системы мочи способны буферировать Н+ при рН менее 6 ед.;
-нейтрализация фосфатами Н+ происходит до рН = 4,0 - 4,5 ед., после достижения которой буферирование достигается только в результате повышения концентрации буфера;
-в норме «титруемая кислотность» составляет от 10 до 40 мэкв Н+ в сутки.
(Левински Н.Г., 1994; Forman J.W., 1986; Hruska K., 1987)

РОЛЬ ПОЧЕК В РЕГУЛЯЦИИ РН
- основное средство секреции Н+ - образование катиона аммония:
Глутамин Глутаминаза I Глутаминовая к-та + Аммиак
Глутаминовая к-та Глутаматдегидрогеназа а-Кетоглутаровая к-та + Аммиак
-с катионом аммония экскретируется 30-50 мэкв Н+ в сутки;
-за счет глутамина образуется 2/3 катиона аммония, 1/4 - поступает из крови, остальное - дезаминирование аминокислот;
-распределение аммиака между кровью, клетками канальцев и канальцевой жидкостью зависит от относительной рН в каждом из этих сегментов;
(Forman J.W., 1986; Hruska K., 1987)
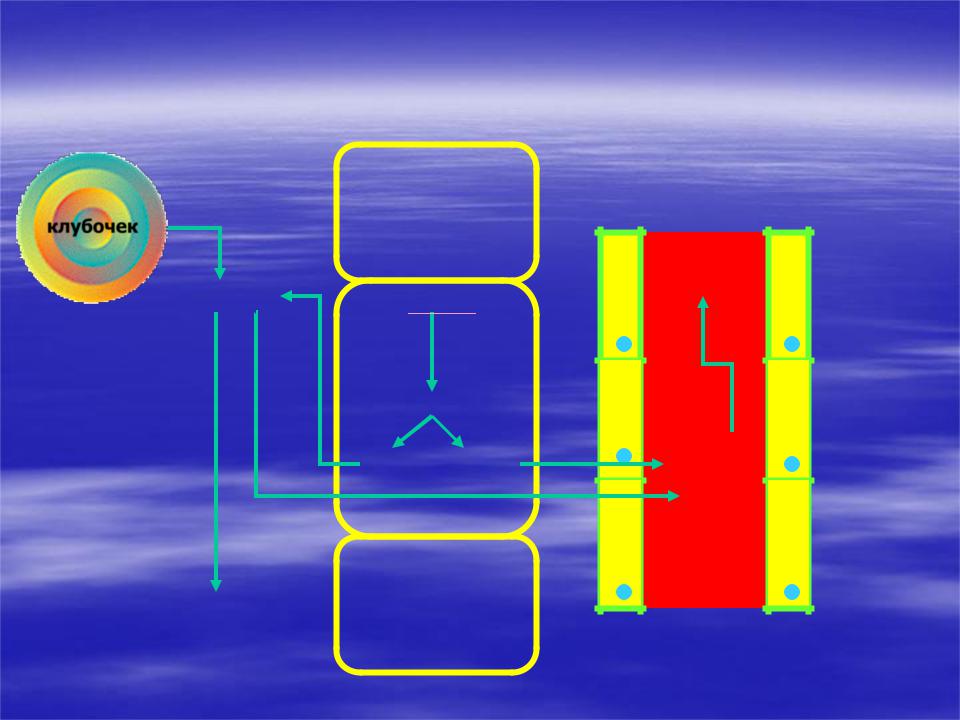
* Регуляция КОС почками (3) Образование свободных слабых органических кислот
просвет канальца
клубочек
CH3COONa
|
|
кровеносный |
|
|
сосуд |
эпителий |
|
|
канальца |
|
|
|
|
NaHCO |
H2O + CO2 |
|
|
|
карбоан- |
|
|
гидраза |
|
|
H2CO |
Na } |
|
3 |
|
|
|
|
Н+ |
HCO3 |
HCO3- |
|
- |
+ |
|
|
|
CH3COOH