

Материалы / 01. Общая микробиология / Тема 1_Введение / Иллюстрированное учебное пособие Общая микробиология
.pdf291
Транспозоны легко перемещаются по хромосоме. При включении в бактериальную хромосому и исключении из нее транспозоны могут вызывать различные виды мутаций (делеции, дупликации, инверсии). Транспозоны не способны к самостоятельной репликации и проявляют свои свойства только в составе другого репликона (бактериальной хромосомы или плазмиды). Транспозоны легко перемещаются с одного репликона на другой (например, с бактериальной хромосомы на плазмиду или ДНК фага). Это способствует широкому распространению транспозонов в бактериальной популяции.
Интегроны представляют собой генетические элементы, которые содержат ген интегразы int, промотор Р и специфические сайты (участки встраивания) att
(рисунок 10.18).
Рисунок 10.18 – Схема строения интегрона.
Такое строение придает интегронам способность захватывать и встраивать в себя небольшие элементы ДНК (мобильные генные кассеты) и экспрессировать присутствующие в их составе гены. Интегроны часто инкорпорируются в плазмиды.
Островки патогенности представляют собой участки ДНК размером от 10000 до 200000п.о. Они содержат до нескольких десятков генов, детерминирующих факторы патогенности бактерий. Участки генома, схожие с островками патогенности, но лишенные генов патогенности, называются геномными или метаболическими островками. Островки патогенности по флангам имеют прямые повторы (DR) или IS-элементы, содержат гены “подвижности” (интегразы, транспозазы) и располагаются вблизи генов тРНК (рисунок 10.19).
Рисунок 10.19 – Схема строения островков патогенности.
Разные участки островков патогенности приобретены бактериями в разное
292
время от разных хозяев в результате горизонтального переноса. Большинство островков патогенности локализовано на хромосоме бактерий (Salmonella), но они также могут находиться в составе плазмид (Shigella) и фаговых ДНК (V. cholerae O1, O139).
10.2.Изменчивость бактерий
Убактерий различают 2 вида изменчивости - фенотипическую и генотипическую. Фенотипическая изменчивость (ненаследственная, модификационная) - это изменение свойств бактерий под влиянием факторов внешней среды без изменения их генетического аппарата. Другими словами фенотипическая изменчивость - это приспособительные реакции бактерий на изменение условий внешней среды. Фенотипическая изменчивость возникает с высокой частотой, а фенотипически измененные бактерии часто реверсирует в исходную форму.
Характерные черты фенотипической изменчивости:
1. Сохранение исходного генотипа и отсутствие передачи по наследству.
2. Причины фенотипической изменчивости - влияние внешних факторов (биологических, химических, физических), блокирующих на время нормальную работу какого-либо фермента.
3. Реверсия к исходным признакам при восстановлении нормальных условий культивирования.
Проявления фенотипической изменчивости:
- изменение морфологических признаков (изменение формы и размера клеток, утрата капсулы, жгутиков);
- изменение культуральных свойств (диссоциация культур, образование R- и S-форм колоний);
- изменение биохимической активности.
Изменчивость морфологических признаков. Под влиянием физических,
химических или биологических агентов у многих микроорганизмов наблюдается изменение формы и размеров клеток. Например, при добавлении стрептомицина к питательной среде клетки сальмонелл значительно удлиняются. При длительном культивировании на питательных средах кишечная палочка принимает кокковидную форму, дифтерийная палочка образует нитевидные или дрожжеподобные формы. Присутствие в питательной среде небольшой концентрации этилового спирта подавляет образование жгутиков у бактерий. Изменение морфологических свойств бактерий представлено на рисунке 10.20.
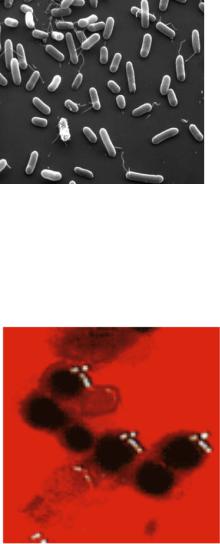
293
Рисунок 10.20 – Изменение размеров и формы бактерий при длительном культивировании на агаре.
Культуральная изменчивость проявляется в образовании колоний разной величины и формы (рисунок 10.21).
Рисунок 10.21 – Изменение формы колоний при длительном выращивании бактерий.
Одной из форм культуральной изменчивости бактерий является диссоциация колоний. При посеве на мясо-пептонный агар в чашки Петри микроорганизмы образуют колонии двух основных типов: S-форма - гладкие колонии (от англ. smooth - гладкий); R-форма - шероховатые колонии (от англ. rough - шероховатый). Феномен образования бактериями разных форм колоний впервые описали в 1917 г. австрийский микробиолог Эдмунд Вейль (1880-1922 гг.) и чехословацкий бактериолог Артур Феликс (1887-1956 гг.). Диссоциация сопровождается изменением биохимических, морфологических, антигенных и патогенных свойств возбудителей.
Свойства клеток из колоний R-формы:
Колонии шероховатые, морщинистые, края колоний изрезанные. Жгутики отсутствуют.
Капсула (слизистый слой) отсутствует. Биохимическая активность снижена.
Вирулентность для лабораторных животных снижена. Антигенная структура неполноценная. Чувствительность к фагу снижена.
294
Клетки полиморфные.
Свойства клеток из колоний S-формы:
Колонии прозрачные, с гладкой блестящей поверхностью, круглые, с ровными краями, выпуклые.
У подвижных видов имеются жгутики.
Хорошо выражена капсула или слизистый слой. Выраженная биохимическая активность. Высокая вирулентность.
Антигенная полноценность. Чувствительность к фагу выражена. Клетки типичной формы и размеров.
Большинство патогенных микробов образует колонии S-формы. Некоторые патогенные бактерии (например, В. anthracis) на обычных питательных средах образуют колонии R-формы, сохраняя при этом вирулентные свойства.
Изменчивость ферментативных (биохимических) свойств обусловлена способностью бактерий синтезировать адаптивные ферменты. Эти ферменты вырабатываются на определенных питательных субстратах и предопределены генотипом. Например, кишечная палочка, растущая на среде без лактозы, не синтезирует разлагающий лактозу фермент, но если пересеять культуру на среду с лактозой, то она начнет вырабатывать этот фермент. Адаптивные ферменты позволяют микробам приспосабливаться к определенным условиям существования.
Генотипическая (наследственная) изменчивость - это изменение свойств бактерий в результате “поломок” генетического материала под влиянием различных факторов. Генотипическая изменчивость сопровождается передачей приобретенных признаков по наследству. Ей присущи низкая частота возникновения и редкая реверсия в исходную форму.
Выделяют 2 формы генотипической изменчивости:
-мутации - изменение последовательности азотистых оснований в ДНК;
-рекомбинации - перенос участков ДНК от бактерии-донора в бактериюреципиент.
Мутации (mutatio - изменение) – это скачкообразные изменения свойств бактерий. Термин “мутация” предложил голландский ботаник Хуго де Фриз как “скачкообразное изменение наследственного признака” у растений. Позднее голландский микробиолог Мартин Бейеринк распространил это понятие на бактерии (рисунок 10.22).
Отечественные микробиологи Г.А. Надсон и Г.С. Филиппов установили, что после воздействия ионизирующим излучением на дрожжевые клетки возникали разнообразные расы, свойства которых воспроизводились в потомстве.
Мутация – это качественное или количественное изменение первичной структуры ДНК, в результате которого утрачивается или изменяется какой-либо признак или группа признаков. Эти изменения могут возникать под влиянием эндогенных или экзогенных факторов. Бактерии с измененными признаками называются мутантами.

295
Рисунок 10.22 – А - Хуго де Фриз (Hugo de Vries, 1848 – 1935 гг.), Б - Мартин Бейеринк (Martinus Willem Beijerinck, 1851 – 1931 гг.).
Классификация мутаций:
1. По происхождению:
-спонтанные (внутренние) мутации;
-индуцированные (внешние) мутации. 2. По протяженности:
-точечные мутации;
-генные мутации;
-хромосомные мутации.
3.По направленности: - прямые мутации; - обратные мутации.
4.По локализации:
-хромосомные мутации;
-плазмидные мутации. 5. По функции:
-летальные мутации;
-сублетальные мутации;
-нейтральные мутации. 6. По фенотипу:
-морфологические мутации;
-функциональные мутации (резистентность, ауксотрофность).
Причинами мутаций могут быть внутренние и внешние факторы. Соответственно этому выделяют спонтанные и индуцированные мутации.
Спонтанные (самопроизвольные) мутации возникают под влиянием внутренних (эндогенных) факторов в результате ошибок ДНК-полимеразы во время репликации ДНК или в результате недостаточности механизмов репарации, то есть исправления ошибок. Частота спонтанных мутаций достигает 1·10-5 - l·10-10. Наиболее распространенным типом спонтанных мутаций у бактерий является ауксотрофность, то есть утрата способности к синтезу какого-либо соединения. Такой мутант называют ауксотрофным по данному соединению. В норме почти все ошибки ДНК-полимеразы исправляются с помощью ферментов репараз. Однако
296
некоторые спонтанные мутации могут закрепиться в потомстве. Например, Л. Пастер в 1881 г. получил вакцину против сибирской язвы, выращивая возбудитель в течение длительного времени при повышенной температуре (42,5ОС). А. Кальмет и Ш. Герен в 1919 г. путем длительных пассажей на питательной среде получили вакцинный штамм микобактерий туберкулеза (вакцину БЦЖ).
Индуцированные (направленные) мутации возникают под действием внешних химических, физических или биологических факторов. Частота возникновения индуцированных мутаций достигает 10-7-10-4, то есть во много раз выше частоты спонтанных мутаций. Физические факторы, вызывающие мутации, называются мутагенными факторами. К ним относятся разные виды излучений. Химические вещества, вызывающие мутации, называются мутагенами. К мутагенам относятся, в частности, соли азотистой кислоты и акридиновые красители. Бактерии с измененными признаками называются мутантами.
Мутации, приводящие к потере функции, называются прямыми. Восстановление исходных свойств называется реверсией. При реверсии исходный генотип может восстанавливаться (обратная мутация) или восстановления исходного генотипа не происходит (супрессорная мутация).
По протяженности (количеству мутировавших генов) выделяют точечные, генные и хромосомные мутации. Точечные мутации затрагивают чаще всего одно основание, генные мутации – один ген, а хромосомная мутация – несколько генов. При некоторых мутациях образуется бессмысленный кодон, не кодирующий ни одну из аминокислот. Такие мутации называют бессмысленными или нонсенс-
мутациями.
Нейтральные мутации фенотипически не проявляются какими-либо изменениями признаков. Условно-летальные (сублетальные) мутации приводят к изменению функциональной активности синтезируемого фермента, но не к утрате синтетической способности. Летальные мутации характеризуются полной утратой способности синтезировать жизненно важные для бактериальной клетки ферменты.
Выделяют следующие типы мутаций.
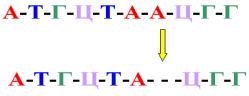
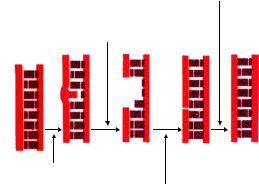
Делеция - это выпадение одного или нескольких пуриновых или пиримидиновых оснований (рисунок 10.22).
Рисунок 10.23 – Схема формирования делеций.
Инсерция – это вставка одного или нескольких пуриновых или пиримидиновых оснований (рисунок 10.24).
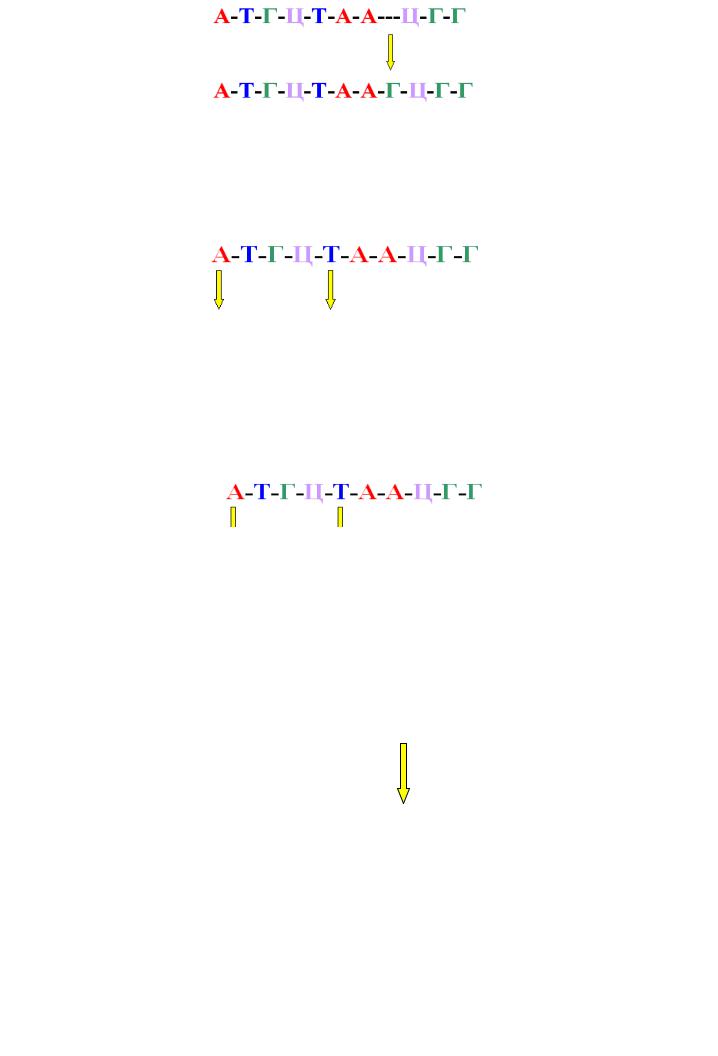
297
Рисунок 10.24 – Схема формирования инсерций.
Транзиция - это замена пурина на пурин или пиримидина на пиримидин (рисунок 10.25).
Рисунок 10.25 – Схема формирования транзиций.
Трансверсия - это замена пурина на пиримидин или пиримидина на пурин (рисунок 10.26).
Рисунок 10.26 - Схема формирования трансверсий.
Дупликация – это удвоение количества пуриновых или пиримидиновых оснований (рисунок 10.27).
А-Т-Г-Ц-Т-А-А-Ц-Г-Г
А-Т-Г-Ц-Т-А-А-Ц-Ц-Г-Г
Рисунок 10.27 – Схема формирования дупликаций.
Дислокация (транслокация) – это перемещение пуриновых или пиримидиновых оснований (рисунок 10.28).
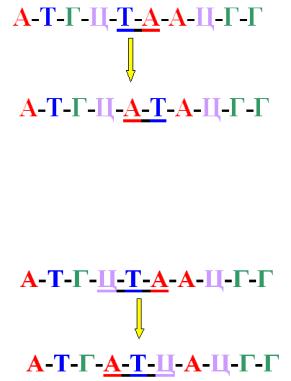
298
Рисунок 10.28 – Схема возникновения дислокаций.
Инверсия – это переворот пуриновых или пиримидиновых оснований относительно друг друга (рисунок 10.29).
Рисунок 10.29 – Схема формирования инверсий.
Некоторые химические вещества значительно повышают частоту мутаций. В частности, аналоги азотистых оснований включаются в молекулу ДНК и вызывают вставку некорректного основания при репликации. Бромурацил, включаясь в ДНК, узнаётся полимеразой как цитозин, в результате чего вместо аденина включается гуанин. Азотистая кислота дезаминирует азотистые основания. Интеркалирующие агенты (акридиновые красители) внедряются между основаниями ДНК и вызывают увеличение расстояния между ними, что приводит к утрате нуклеотидов или включению дополнительной пары нуклеотидов.
В мутантных клетках существуют механизмы, способные полностью или частично восстанавливать исходную структуру ДНК. Способность исправлять повреждения в молекуле ДНК называется репарацией. Репарация осуществляется специальными ферментными системами клетки. Клеточные ферменты репарации выполняют строго определенные функции:
-ферменты, узнающие измененные участки ДНК и осуществляющие разрыв цепи рядом с поврежденным участком;
-ферменты, удаляющие поврежденные участки ДНК;
-ферменты, синтезирующие участок ДНК взамен удаленного;
-ферменты, восстанавливающие непрерывность цепи ДНК.
У бактерий выделяют 3 основных механизма репарации:
-прямая репарация (исходная структура нуклеотидов восстанавливается в одну стадию);
-эксцизионная репарация (удаление поврежденных оснований с последующим восстановлением исходной структуры ДНК);
-пострепликативная репарация (после репликации образуется ДНК, имеющая одноцепочечные бреши, которые заполняются в процессе рекомбинации).
299
К прямой репарации относится световая репарация или фотореактивация (исправление повреждения ДНК под действием УФ-лучей). Световая репарация осуществляется с помощью таких ферментов как фотолиаза, метилтрансфераза, инсертаза, гликозилаза. Под действием конкретного фермента безошибочно восстанавливается исходная структура ДНК.
При эксцизионной репарации место повреждения распознается эндонуклеазой, которая расщепляет ДНК вблизи дефекта. Затем поврежденный фрагмент удаляется, а в образовавшуюся брешь при помощи ДНК-полимеразы встраиваются отсутствующие нуклеотиды (ресинтез нуклеотидной цепи на основе неповрежденной матрицы). Схема репарации ДНК представлена на рисунке 10.30.
Сшивка разрыва ДНК-лигазой
Удаление поврежденного участка
Повреждение
Восполнение дефекта ДНК-полимеразой
Рисунок 10.30 – Схема репарации ДНК.
Рекомбинация - это изменчивость, обусловленная переносом участка ДНК от бактерии-донора в бактерию-реципиент. У бактерий выделяют гомологичная и сайт-специфическую рекомбинация. Гомологичная рекомбинация происходит под контролем генов REC-системы (гены recA, B, C, D). Рекомбинируют ДНК с высокой степенью гомологии. Сайт-специфическая рекомбинация не зависит от REC-
системы и гомологии ДНК. Наблюдается при встраивании плазмид и умеренных фагов в определенные участки бактериальной хромосомы за счет IS-элементов.
Рекомбинация у бактерий происходит при реализации следующих механизмов:
-трансформация;
-конъюгация;
-трансдукция.
Трансформация (преобразование, перестройка) - это изменение свойств бактерий в результате поступления (поглощения, искусственного введения) в клетку-реципиент свободного фрагмента ДНК, изолированного из клетки-донора. Трансформация возможна как in vivo, так и in vitro. Явление трансформации открыл в опытах на пневмококках в 1928 г. английский генетик Ф. Гриффит (рисунок
10.31).
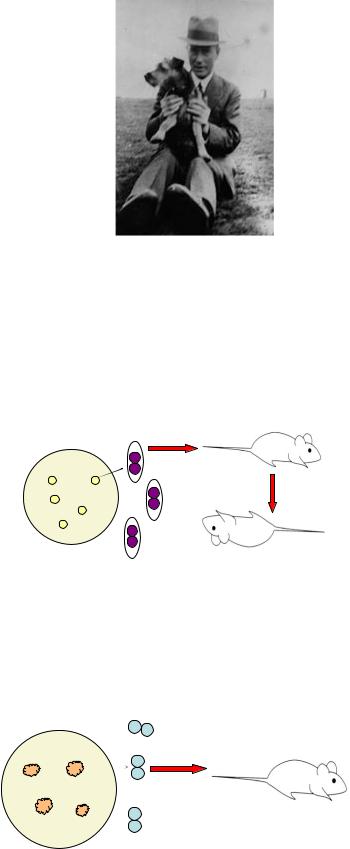
300
Рисунок 10.31 – Фредерик Гриффит (Frederick Griffith, 1879 - 1941 гг.).
Эксперимент Ф. Гриффита заключался в следующем. Пневмококки на питательном агаре могут образовывать колонии S- и R-формы. Пневмококки, синтезирующие капсулу, формируют на плотной питательной среде гладкие колонии S-формы. При введении мышам капсульных пневмококков отмечается гибель животных (рисунок 10.32).
Рисунок 10.32 – Патогенность для мышей капсульных пневмококков.
Бескапсульные пневмококки на питательном агаре образуют шероховатые колонии R-формы. При введении мышам бескапсульных клеток животные остаются живыми (рисунок 10.33).
Рисунок 10.33 – Патогенность для мышей бескапсульных пневмококков.
При кипячении бактериальной суспензии капсульные клетки погибают и при введении мышам гибели животных не отмечается (рисунок 10.34).