

Bykov-_gistologia_obschaya
.pdfОСОБЫЕ ТИПЫ ГЛАДКИХ МИОЦИТОВ
Помимо описанной выше гладкой мышечной ткани, в организме человека имеются несколько особых типов гладких миоцитов, которые кратко охарактеризованы ниже. К ним относятся, в первую очередь, миоэпителиальные и мионейральные клетки, которые по своему происхождению отличаются от основного типа гладкой мышечной ткани, развивающегося из мезенхимы, и поэтому иногда описываются как отдельные гистогенетические типы мышечных тканей. В группу гладких миоцитов с особыми структурнофункциональными свойствами относят также эндокринные гладкие миоциты и миофибробласты.
Миоэпителиальные клетки происходят из эктодермы (а также, возможно, частично, из прехордальной пластинки) и представляют собой видоизмененные эпителиальные клетки. Они не обладают исчерченностью (относятся к гладким) и входят в состав концевых отделов (частично - мелких выводных протоков) потовых, молочных, слезных и слюнных желез, а также желез трахеи и пищевода. С железистыми клетками миоэпителиальные клетки связаны десмосомами, снаружи они покрыты базальной мембраной. Сокращаясь, миоэпителиальные клетки способствуют выведению секрета из концевых отделов и выводных протоков.
Рис. 13-17. Миоэпителиальные клетки в концевом отделе экзокринной железы. 1 - вид с поверхности (базальная мембрана удалена), 2 - вид на разрезе. МЭК - миоэпителиальные клетки, ЖК - железистые клетки, БМ - базальная мембрана.
Форма миоэпителиальных клеток в концевых отделах - отростчатая,
звездчатая. Здесь эти клетки получили также название корзинчатых, так как в совокупности они образуют своеобразную "корзинку", охватывающую железистые клетки концевого отдела (рис. 13-17). В протоках желез чаще встречаются клетки веретеновидной формы, циркулярно охватывающие эпителиальную трубочку. Ядро занимает
- 451 -
в клетке центральное положение; в цитоплазме, преимущественно в отростках, выявляются миофиламенты, образующие сократительный аппарат миоэпителиальных клеток, а также многочисленные цитокератиновые промежуточные филаменты, свойственные эпителиальным клеткам. Иммуногистохимическими методами в них выявляется также и десмин - белок промежуточных филаментов, характерный для мышечных тканей.
Мионейральные клетки имеют нейральное происхождение - они развиваются из клеток наружного слоя глазного бокала, являются гладкими и образуют мышцы радужки глаза (суживающую и расширяющую зрачок).
Мышца, суживающая зрачок, состоит из клеток, которые имеют веретеновидную форму и, располагаясь циркулярно, образуют мышечное кольцо у дистального (свободного) края радужки. По строению и функции они сходны с гладкими миоцита ми мезенхимного происхождения.
Мышца, расширяющая зрачок, образована расположенными радиально в радужке отростками клеток, ядросодержащие части ("тела") которых находятся между задним пигментным эпителием и стромой радужки. Тела этих мышечных клеток, в отличие от отростков, заполнены гранулами пигмента (отчего их называют также миопигментоцитами) и связаны десмосомами с клетками заднего эпителия. Многие авторы описывают эти клетки как видоизмененный второй слой пигментного эпителия радужки и поэтому считают их миоэпителиальными.
Эндокринные гладкие миоциты (юкстагломерулярные, эпителиоидные, зернистые клетки) являются видоизмененными гладкими миоцитами, которые представляют собой основной компонент юкстагломерулярного аппарата почек. Они входят в состав стенки артериол почечного тельца и характеризуются редуцированным сократительным аппаратом при выраженном развитии синтетического аппарата. Продуцируемый этими клетками фермент ренин накапливается в их цитоплазме в виде покрытых мембраной гранул, содержимое которых выводится в кровь механизмом экзоцитоза.
Миофибробласты также относятся к клеткам с сократительной функцией. Они представляют собой видоизмененные фибробласты - клетки соединительной ткани, участвующие в выработке волокон и основного вещества, но одновременно обладающие выраженными сократительными свойствами (см. главу 10). Вариантами миофибробластов, по-видимому, являются миоидные клетки, входящие в состав стенки извитого семенного канальца яичка и наружного слоя теки фолликула яичника.
- 452 -

Глава 14
НЕРВНАЯ ТКАНЬ
Нервная ткань является функционально ведущей тканью нервной системы; она состоит из нейронов (нейроцитов, собственно нервных клеток), обладающих способностью к выработке и проведению нервных импульсов, и клеток нейроглии, выполняющей ряд вспомогательных функций (опорную, трофическую, барьерную, защитную и др.) и обеспечивающей деятельность нейронов. Нейроны и нейроглия (за исключением одной из ее разновидностей - микроглии) являются производными нейрального зачатка.
ГИСТОГЕНЕЗ НЕРВНОЙ ТКАНИ
Нервная пластинка представляет собой нейральный зачаток - источник развития нервной ткани в эмбриогенезе. У 16-дневного зародыша человека она имеет вид удлиненного дорсального утолщения эктодермы, лежащего над хордой. Детерминация материала нервной пластинки происходит в результате второй фазы гаструляции под индуцирующим влиянием хордомезодермального зачатка. При обособлении нейрального зачатка (нейруляции) выделяются три его компонента: нервная трубка, нервный гребень и нейральные плакоды.
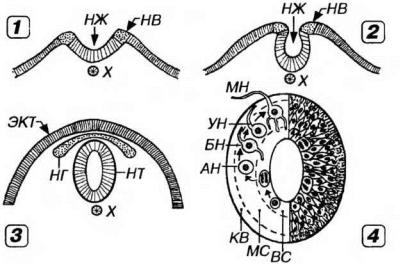
Нервная трубка. В процессе выделения и обособления нервного зачатка (18-21-й дни развития эмбриона человека) нервная пластинка прогибается, превращаясь сначала в нервный желобок (с приподнятыми краями - нервными валиками), который затем (22-й день) замыкается в нервную трубку и обособляется от эктодермы (рис. 14-1).
Производными нервной трубки являются нейроны и глия органов центральной нервной системы (ЦНС) - головного и спинного мозга, а также ряд структур периферической нервной системы (ПНС).
Нервный гребень. При смыкании нервной трубки в области нервных валиков между ней и кожной эктодермой с обеих сторон выделяются скопления клеток, образующие нервный гребень, называемый также ганглиозной пластинкой (см. рис. 14-1). Клетки нервного гребня утрачивают взаимные адгезивные связи и осуществляют миграцию в вентральном и латеральном направлениях в виде нескольких рассеивающихся потоков, которые дают многочисленные производные. Ход последующей дифференцировки клеток нервного гребня, в соответствии
- 453 -
с одними взглядами, запрограммирован еще до их миграции, согласно другим - определяется их микроокружением в течение миграции и в ее конечном участке, а также временем миграции.
Рис. 14-1. Гистогенез нервной ткани: нейруляция (1-3) и строение нервной трубки (4). В ходе нейруляции прогибание нервной пластинки (1-2) приводит к образованию нервного желобка (НЖ) с приподнятыми краями - нервными валиками (НВ). 3 - замыкание НЖ в нервную трубку (НТ) обусловливает выделение материала НВ в нервный гребень (НГ) и обособление нервного зачатка от кожной эктодермы (ЭКТ). X - хорда. Стенка НТ у эмбриона на 3-4-й нед. развития состоит из трех слоев (изнутри кнаружи): вентрикулярного (ВС), содержащего камбиальные элементы и митотически делящиеся клетки, мантийного (МС) образованного клетками, мигрирующими из ВС и дифференцирующимися в нейробласты и спонгиобласты, и краевой вуали (КВ), которая содержит отростки клеток, расположенных в МС и ВС. В МС происходит последовательное превращение нейробластов из аполярных (АН) в биполярные (БН), униполярные (УН) и мультиполярные (МН), которые постепенно дифференцируются в зрелые нейроны.
Производными нервного гребня являются нейроны и глия спинальных, вегетативных ганглиев и ганглиев некоторых черепно-мозговых нервов, леммоциты, клетки мозгового вещества надпочечников, диффузной эндокринной системы, паутинной и мягкой мозговой оболочек, пигментные клетки (меланоциты). В краниальной части он служит также источником эктомезенхимы, которая дает начало части скелетных и волокнистых соединительных тканей области головы и шеи, аорты и сердца.
- 454 -
Плакоды (от греч. plax - пластинка) - утолщённые участки эктодермы в краниальной части зародыша по краям от нервной трубки, клетки которых обладают нейральной детерминацией, но не участвуют в образовании нервной трубки и нервного гребня.
Производными плакод являются некоторые клетки органов чувств - слуха, равновесия, вкуса (рецепторные, поддерживающие и выстилающие канальцы) и зрения (эпителий хрусталика).
Замыкание нервной трубки начинается в шейном отделе в области появления первых сомитов, распространяясь в дальнейшем краниально и каудально. Открытые края нервной трубки (краниальный и каудальный нейропоры) замыкаются на 24-й и 26-й дни внутриутробного развития, соответственно. Из расширяющегося краниального отдела нервной трубки, дающего начало трем первичным мозговым пузырям, формируется головной мозг, из остальной ее части образуется спинной мозг.
Стенка нервной трубки на ранних стадиях развития состоит из одного слоя клеток призматической формы, которые интенсивно делятся и мигрируют от ее просвета, в результате чего на 3-4-й нед. в ней можно выделить три слоя (изнутри кнаружи):
1)вентрикулярный (матричный, эпендимный) слой содержит камбиальные элементы и митотически делящиеся метки. Часть клеток, образующих внутреннюю выстилку нервной трубки, дает начало эпендимной глии;
2)мантийный (плащевой) слой пополняется, в основном, за счет миграции клеток из эпендимного слоя, которые дифференцируются в нейробласты (дают начало нейронам) или спонгиобласты (глиобласты), дающие начало астроцитарной глии и олигодендроглии. Один из видов глиобластов преобразуется в радиальные глиальные клетки, которые протягиваются через всю стенку нервной трубки и служат направляющими элементами для миграции нейробластов. В дальнейшем радиальные глиальные клетки дифференцируются
вастроциты.
3)краевая вуаль содержит отростки клеток, расположенных в двух более глубоких слоях.
Нейробласты сначала не имеют отростков (аполярные нейробласты), затем на противоположных концах их тел формируются отростки (клетки превращаются в биполярные нейробласты). Один из отростков подвергается обратному развитию (клетки преобразуются в униполярные нейробласты), на месте утраченного отростка в дальнейшем появляется несколько новых (дендритов), а нейробласты становятся мультиполярными, постепенно дифференцируясь в зрелые нейроны, которые утрачи-
- 455 -
вают способность к делению. Дифференцировка нейробласта в нейрон сопровождается накоплением в его цитоплазме цистерн грЭПС, увеличением объема комплекса Гольджи, накоплением элементов цитоскелета.
Рост аксона нейрона происходит со скоростью около 1 мм/сут.; он продвигается в тканях амебоидными движениями к иннервируемому им органу (органу-мишени), очевидно вследствие тропизма к выделяемым этим органом веществам. Рост ускоряется под действием фактора роста нервов (ФРН). На конце растущего аксона имеется расширение (конус роста), состоящее из центральной уплощенной части, от которой отходят тонкие (0.1-0.2 мкм) длинные (до 50 мкм) отростки (микро-шипики, филоподии), содержащие многочисленные актиновые микрофиламенты и непрерывно меняющие свою форму и длину. Конус роста обеспечивает направленный рост аксона благодаря распознаванию контактных (адгезивных) и дистантных (гуморальных) химических сигналов. Рост аксона завершается его прикреплением к органумишени. За первым аксоном, вступающим в связь с органом-мишенью (аксоном-пионером), устремляются другие, формируя в дальнейшем тракты в ЦНС и нервы в ПНС.
Гибель нейронов в эмбриональном развитии происходит в значительных масштабах, охватывая 40-85% клеток в различных участках нервной системы (в частности, более половины двигательных нейронов) и осуществляется механизмом апоптоза. Причина этого явления, как предполагают, заключается в том, что нейроны, не установившие связи с клетками органа-мишени, не получают необходимых для поддержания их жизнедеятельности трофических факторов, выделяемых этим органом и поглощаемых их аксонами. Гибель нейронов может происходить и вследствие избыточной иннервации органовмишеней; возможно, при этом устраняются также и неправильно сформировавшиеся связи.
НЕЙРОНЫ
ОБЩИЕ СВЕДЕНИЯ
Нейроны (нейроциты, собственно нервные клетки) - клетки различных размеров (которые варьируют от самых мелких в организме - у нейронов с диаметром тела 4-5 мкм - до наиболее крупных с диаметром тела около 140 мкм). Их общее количество в нервной системе человека превышает 100 млрд. (1011), а по некоторым оценкам достигает одного триллиона (1012). К рождению нейроны утрачивают способность
- 456 -
к делению, поэтому в течение постнатальной жизни их количество не увеличивается, а, напротив, в силу естественной убыли клеток, постепенно снижается.
Гибель нейронов в физиологических условиях у взрослого человека
сравнительно невелика и осуществляется механизмом апоптоза. Избыточной потере нейронов препятствует их относительно высокая устойчивость к развитию апоптоза, характерная для всех необновляемых клеток. Гибель нейронов значительно ускоряется в старости, приводя к потере 20-40% клеток в некоторых участках головного мозга.
Гибель нейронов при дегенеративных заболеваниях нервной системы
(болезнях Альцгеймера, Гентингтона, Крейцфельда-Якоба, паркинсонизме, боковом амиотрофическом склерозе и др.) осуществляется вследствие ненормально высокой активности апоптоза, что приводит к резкому снижению их содержания в определенных участках ЦНС. Развитие неврологических нарушений, которые выявляются у 90% больных СПИДом, связано с потерей 40-50% нейронов в коре головного мозга, которые также погибают путем апоптоза.
ФУНКЦИОНАЛЬНАЯ МОРФОЛОГИЯ НЕЙРОНА
Нейрон состоит из клеточного тела (перикариона) и отростков, обеспечивающих проведение нервных импульсов - дендритов, приносящих импульсы к телу нейрона, и аксона (нейрита), несущего импульсы от тела нейрона (рис. 14-2 и 14-3).
Тело нейрона (перикарион) включает ядро и окружающую его цитоплазму (за исключением входящей в состав отростков). Перикарион содержит синтетический аппарат нейрона, а его плазмолемма осуществляет рецепторные функции, так как на ней находятся многочисленные нервные окончания (синапсы), несущие возбуждающие и тормозные сигналы от других нейронов.
Ядро нейрона - обычно одно, крупное, округлое, светлое, с мелкодисперсным хроматином (преобладанием эухроматина), одним, иногда 2- 3 крупными ядрышками. Эти особенности отражают высокую активность процессов транскрипции в ядре нейрона. Около ядрышка в нейронах у лиц женского пола часто выявляется тельце Барра - крупная глыбка хроматина, содержащая конденсированную Х-хромосому (особенно заметна в клетках коры полушарий большого мозга и симпатических нервных узлов).
- 457 -
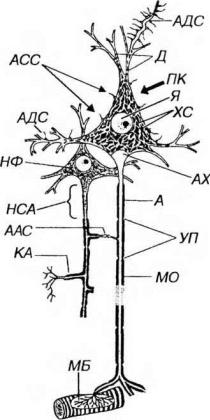
Рис. 14-2. Строение мультиполярного нейрона (по Rohen J.W., Lutjen-Drecoll E. 1982). ПК - перикарион, Я - ядро с ядрышком, ХС - хроматофильная субстанция, НФ - нейрофибриллы (агрегаты элементов цитосклета), Д - дендриты. А - аксон, НСА - начальный сегмент аксона, АХ - аксонный холмик, КА - коллатерали аксона, МО - миелиновая оболочка, УП - узловые перехваты, МБ - моторная бляшка (двигательное нервное окончание на волокне поперечнополосатой мышцы). Синапсы (С): АДС - аксо-дендритический, АСС - аксо-соматический, ААС - аксо-аксональный.
Цитоплазма нейрона богата органеллами и окружена плазмолеммой, которая обладает способностью к проведению нервного импульса (распространению деполяризации) вследствие локального тока Na+ в цитоплазму и К+ из нее через потенциал-зависимые мембранные ионные каналы. Плазмолемма содержит Na+-K+ насосы, которые поддерживают необходимые градиенты ионов.
- 458 -
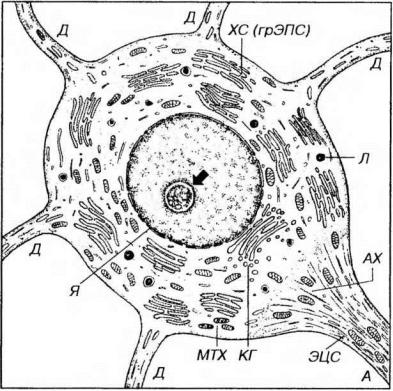
Рис. 14-3. Ультраструктурная организация нейрона. Я - ядро (ядрышко показано стрелкой), ХС - хроматофильная субстанция, ЭЦС - элементы цитоскелета (нейротрубочки, нейрофиламенты), МТХ - митохондрии, КГ - комплекс Гольджи, Л -лизосомы, Д - дендриты, А - аксон, АХ - аксонный холмик.
грЭПС хорошо развита, ее цистерны часто образуют отдельные комплексы из параллельно лежащих уплощенных анастомозирующих элементов, которые на светооптическом уровне при окраске анилиновыми красителями имеют вид базофильных глыбок, в совокупности получивших название хроматофильной субстанции (вещества, или телец Ниссля, тигроидного вещества, тигроида). Характер распределения и размеры комплексов цистерн грЭПС (хроматофильной субстанции) варьируют в отдельных типах нейронов (наиболее крупные обнаруживаются в мотонейронах) и зависят от их функционального состояния. При длительном раздражении или повреждении нейрона комплексы цистерн грЭПС распадаются на отдельные элементы, что на светооптическом уровне проявляется исчезновением телец Ниссля (хроматолиз, тигролиз).
- 459 -
аЭПС образована трехмерной сетью анастомозирующих цистерн и трубочек, участвующих в синтетических процессах и внутриклеточном транспорте веществ.
Комплекс Гольджи хорошо развит (впервые описан именно в нейронах) и состоит из множественных диктиосом, расположенных обычно вокруг ядра.
Митохондрии - очень многочисленны и обеспечивают высокие энергетические потребности нейрона, связанные со значительной активностью синтетических процессов, проведением нервных импульсов, деятельностью ионных насосов. Они обычно имеют палочковидную форму и характеризуются быстрым изнашиванием и обновлением (коротким жизненным циклом).
Лизосомальный аппарат (аппарат внутриклеточного переваривания)
обладает высокой активностью и представлен эндосомами и многочисленными лизосомами различных размеров. Интенсивные процессы аутофагии обеспечивают постоянное обновление компонентов цитоплазмы нейрона. При дефектах некоторых лизосомальных ферментов в цитопламзе нейронов накапливаются непереваренные продукты, что нарушает их функции и вызывает болезни накопления, например, ганглиозидоз (болезнь Тэй-Закса).
Цитоскелет нейронов хорошо развит и представлен всеми элементами - микротрубочками (нейротрубочками), микрофиламентами и промежуточными филаментами (нейрофиламентами). Они образуют трехмерную опорносократительную сеть, играющую важную роль в поддержании формы этих клеток и, в особенности, их длинного отростка - аксона. Многочисленные промежуточные филаменты нейрофиламенты) связаны друг с другом и с нейротрубочками поперечными мостиками; при фиксации они склеиваются в пучки, которые окрашиваются солями серебра. Такие образования (фактически являющиеся артефактами) на светооптическом уровне описаны под названием нейрофибрилл - нитей толщиной 0.5-3 мкм, образующих сеть в перикарионе. Микротрубочки (нейротрубочки) и микрофиламенты имеют такое же строение, как и в других клетках. Клеточный центр присутствует во всех нейронах, его главная функция - сборка микротрубочек.
Включения в цитоплазме нейрона представлены липидными каплями, гранулами липофусцина (пигмента старения, или изнашивания, который, однако, выявляется даже в нейронах плодов), (нейро)меланина - в нейронах черной субстанции (substantia nigra) и голубого пятна (locus coeruleus).
Дендриты проводят импульсы к телу нейрона, получая сигналы от других нейронов через многочисленные межнейронные контакты (аксодендритические синапсы), расположенные на них в области осо-
- 460 -