

Пищевая Биохимия / Рогожин В.В., Рогожина Т.В. Биохимия сельскохозяйственной продукции
.pdf14.4. Биосинтез РНК
дочерние цепи ДНК. Наличие фрагментов Оказаки указывает на то, что синтез ДНК происходит прерывисто, отдельными участками.
Таким образом, в процессе биосинтеза белка можно выделить три стадии (инициация, элонгация и терминация). На первой стадии (инициация) в процессе участвуют белки и ферменты, осуществляющие ферментативный биосинтез на матрице ДНК затравочного олигорибонуклеотида (праймера) со свободной гидроксильной группой у С-3′рибозы. При этом к цепям ДНК последовательно присоединяются ДНК-раскручивающие и ДНК-связывающие белки, а затем комплексы ДНК-полимераз и праймаз (см. рис. 14.6). Инициация служит единственной стадией репликации ДНК, которая строго контролируется действием регуляторных систем клетки.
На второй стадии (элонгация) проявляют действие ферменты, которые осуществляют синтез лидирующей и отстающей полинуклеотидных цепей на обеих материнских цепях ДНК. Синтез лидирующей цепи начинается
ссинтеза праймера (при участии праймазы) у точки начала репликации, затем к праймеру присоединяются дезоксирибонуклеотиды под действием ДНК-полимеразы III; далее синтез протекает непрерывно, следуя шагу репликационной вилки. Синтез отстающей цепи протекает в направлении, обратном движению репликационной вилки и начинается фрагментарно. Фрагменты всякий раз синтезируются раздельно, начиная
ссинтеза праймера, который может переноситься с готового фрагмента при помощи одного из белковых факторов репликации в точку старта биосинтеза последующего фрагмента противоположно направлению синтеза фрагментов. Завершается процесс синтеза ДНК (терминация) отделением олигорибонуклеотидных праймеров, объединением отдельных фрагментов ДНК при помощи ДНК-лигаз и формированием дочерней цепи ДНК. Точность репликации ДНК чрезвычайно высока, возможна одна ошибка на 1010 трансферазных реакций, однако подобная ошибка обычно легко исправляется за счет процессов репарации.
14.4. БИОСИНТЕЗ РНК
Процесс синтеза преРНК, путем переписывания информации с ДНК, называется транскрипцией (от англ. transcription — переписывание). Транскрипция осуществляется с помощью различных РНК-полимераз. В эукариотической клетке присутствует четыре вида РНК-полимераз: РНК-полимеразы I, II, III — которые располагаются в ядре клетки и одна — в митохондриях. РНК-полимераза I отвечает за синтез рибосомных 18S, 28S и 5,8S РНК. Рибосомальная 5S РНК и транспортные РНК синтезируются РНК-полимеразой III, а РНК-полимераза II осуществляет синтез предшественников мРНК. В структуре ДНК выявляются особые
243

Глава 14. Метаболизм нуклеиновых кислот
регуляторные элементы, названные энхансерами, в присутствии которых резко возрастает скорость транскрипции; при этом энхансеры проявляют активность независимо от ориентации и положения относительно гена. То есть они могут быть перед геном, внутри или за ним. Индукция генов может осуществляться с помощью фитогормонов, которые в составе комплекса с белком-рецептором способны связываться с регуляторной последовательностью ДНК, что сопровождается активированием процесса транскрипции.
Ингибирование процесса транскрипции осуществляется белком-реп- рессором за счет его специфического связывания с регуляторным участком в структуре ДНК или РНК. Этот процесс называется репрессией генов.
14.5. ПРОЦЕССИНГ И СПЛАЙСИНГ
Процесс формирования зрелых молекул РНК транспортной (тРНК), матричной (мРНК), рибосомальной (рРНК) из неактивного предшественника (преРНК) в эукариотической клетке называется процессингом, или посттранскрипционной модификацией. В результате процессинга происходят следующие действия: отрезание «лишних» концевых последовательностей; расщепление длинных первичных транскриптов; вырезание их них участков, транскрибированных с интронов; добавление нуклеозидов к 3′-концу транскрипта; добавление нуклеотидов к 5′-концу транскрипта; модификация оснований в транскрипте. При этом конечным этапом процессинга является метилирование мРНК, в результате которого на каждые 400 остатков аденинов приходится один остаток 6-метиладе- нина. Процессинг завершается тем, что функционально активная мРНК в виде нуклеопротеидного комплекса, в составе которого набор белковинформоферов, покидает ядро клетки через поры в ядерной мембране, поступая в цитоплазму для трансляции.
Конечным этапом процессинга является процесс, называемый сплайсингом, в течение которого происходит формирование функционально активной мРНК из преРНК. В процессе сплайсинга происходит вырезание из первичного транскрипта преРНК интронных (вставочных) участков, с последующим соединением участков, которые называются экзонами.
При этом экзонами являются участки в структуре ДНК и преРНК, несущие генетическую информацию о структуре белка и чередующиеся с интронами. В процессе сплайсинга интронные участки вырезаются, а экзоны сшиваются между собой, образуя функционально активную мРНК. Считывание информации с мРНК происходит на рибосоме, обеспечивая упорядоченный синтез полинуклеотидной цепочки белка. Сплайсинг протекает на сплайсосоме, которая является специализированной внут-
244

14.6. Функции нуклеиновых кислот
риядерной многокомпонентной структурой, включающей десятки белков и набор малых ядерных РНК.
Синтезированная в процессе транскрипции РНК называется матричной (информационной) РНК и содержит точную копию генетической информации, закодированной в определенном участке ДНК. При этом информация в дальнейшем реализуется в упорядоченном синтезе полипептидной цепочки, в которой определяется количественный и качественный состав аминокислот, а также последовательность их связывания
впервичной структуре белка. Эта информация закодирована в мРНК
ввиде кодонов (триплетов), которые представляют три последовательно связанных между собой остатка нуклеотидов. Вся совокупность триплетов, передающих информацию о 20 аминокислотных остатках, которые будут упорядоченно включены в структуру полипептидной цепи, называется
генетическим кодом.
14.6. ФУНКЦИИ НУКЛЕИНОВЫХ КИСЛОТ
Дезоксирибонуклеиновая кислота. Основные места нахождения ДНК в растительных клетках — ядро, митохондрии и хлоропласты. На ДНК возлагаются следующие функции: 1) хранение и участие в передаче информации о всех белках, которые должны быть синтезированы в клетке; 2) в ДНК содержится информация об индивидуальных свойствах и проявлениях признаков растительного организма; 3) является основным элементом в передаче информации при воспроизводстве растительных организмов; 4) участвует в передаче информации во время деления клеток; 5) определяет индивидуальные свойства клеток различных органов и тканей растений; 6) содержит информацию о структуре клетки и клеточных органеллах; 7) через управляемый синтез белков (ферментов) регулирует протекание ферментативных реакций в клетке, метаболических и функциональных процессах; 8) является основным элементом управления клеток; 9) определяет время жизни соматической клетки.
Рибонуклеиновые кислоты. тРНК выполняет следующие функции: 1) обеспечивает специфическое связывание с аминоацил-тРНК-синтетазой и последующее связывание аминокислот согласно информации антикодона; 2) осуществляет перенос аминокислоты к месту синтеза белка; 3) участвует в процессе распознавания кодона мРНК и синтезе полипептидной цепочки на рибосомах, т. е. обеспечивает специфическое присоединение каждой аминокислоты к растущей полипептидной цепочке.
мРНК участвует в следующих процессах: 1) инициирует формирование функционально активной структуры рибосомы; 2) согласно данных кодона производит избирательное связывание аминоацил-тРНК в ами-
245

Глава 14. Метаболизм нуклеиновых кислот
ноацильном участке рибосомы; 3) обеспечивает избирательное связывание аминокислот в полипептидной цепочке согласно информации, записанной в кодоне.
рРНК осуществляет контроль за процессом синтеза полипептидной цепочки на рибосоме.
ВОПРОСЫ И ЗАДАНИЯ ДЛЯ САМОКОНТРОЛЯ
1. Расскажите о процессах синтеза пуриновых и пиримидиновых нуклеотидов
вклетках растений. 2. Назовите основные ферменты, принимающие участие в реакциях синтеза пуриновых оснований. 3. Какие ферменты участвуют в расщеплении нуклеиновых кислот? 4. Опишите строение ксантиноксидазы. 5. Расскажите о процессе синтеза ДНК в клетках растений. 6. Опишите механизм образования фрагментов Оказаки. 7. Расскажите о процессе биосинтеза РНК. 8. Укажите отличительные особенности в механизмах процессинга и сплайсинга. 9. Назовите основные функции дезоксирибонуклеиновой кислоты. 10. Укажите на различия
вфункционировании различных РНК.
ГЛАВА 15
МЕТАБОЛИЗМ БЕЛКОВ
Втканях растений постоянно протекают процессы синтеза белков
иих распад. Причем скорость этих процессов зависит от вида, возраста растения, условий его произрастания, а также жизненного цикла клеток. В период активного роста растений процессы синтеза белков преобладают над распадом. Однако по мере созревания растительного организма скорость синтетических процессов начинает понижаться. В молодых органах процессы биосинтеза протекают быстрее, чем в стареющих органах.
Фотосинтез оказывает влияние на биосинтез белков. Высокая фотосинтетическая активность растительных тканей всегда сопровождается возрастанием скорости биосинтеза белков, тогда как длительное пребывание растения в темноте способствует повышению скорости распада белков в клетках растений. Каждый вид растения характеризуется специфичным набором белков, что определяет видовые особенности растительного организма и указывает на индивидуальность информации, хранящейся в ядерной ДНК растительного организма.
Следует отметить, что механизмы биосинтеза белков у всех живых организмов практически одинаковы и состоят из нескольких этапов. На начальном этапе образуются функционально активные РНК (матричная, транспортная и рибосомальная), а затем происходит синтез полипептидной цепочки на рибосоме. Заканчивается процесс биосинтеза белка формированием функционально активного белка, имеющего третичную
ичетвертичную структуру.
15.1.БИОСИНТЕЗ ПОЛИПЕПТИДНОЙ ЦЕПОЧКИ БЕЛКА НА РИБОСОМЕ
Процесс перевода информации, заложенной в последовательности нуклеотидов мРНК, в последовательность аминокислотных остатков полипептидной цепи называется трансляция (от англ. translation — перевод).
247

Глава 15. Метаболизм белков
На включение в белок каждой аминокислоты расходуется энергия четырех высокоэнергетических связей (одной молекулы АТФ на стадии синтеза аминоацил-тРНК и трех молекул ГТФ — на стадиях связывания амино- ацил-тРНК и трансляции). В процессе трансляции принимают участие все три вида РНК. Причем, если в матричной РНК информация записана в виде кодонов (триплетов), т. е. трех последовательно связанных нуклеотидов, то для считывания этой информации используется транспортная РНК. Тогда как проверяет правильность считывания закодированной информации рибосомальная РНК. При этом транспортная РНК представляет из себя небольшие последовательности (75…90) мононуклеотидов, содержащие антикодон из трех последовательно связанных между собой нуклеотидов, комплиментарный кодону для аминокислоты в информационной РНК, расположенный в тРНК в месте локализации антикодоновой петли, недалеко от вариабельной петли (см. рис. 4.5). Функция тРНК состоит в том, чтобы транспортировать аминокислоты к рибосоме и вставлять их в определенные участки полипептидной цепи при ее биосинтезе, переводя последовательность нуклеотидов в кодоне мРНК в последовательность аминокислотных остатков первичной структуры белка (рис. 15.1).
Генетический код для аминокислот является вырожденным, т. к. некоторые аминокислоты закодированы 2…6 кодонами (табл. 15.1). Исключение представляют триптофан и метионин, имеющие по одному кодону. Всего имеется 64 кодона, три из которых не кодируют никакой аминокислоты; УАГ, УАА и УГА обозначают конец матрицы: на этих триплетах обрывается дальнейшее наращивание пептидной цепи — терминирующие триплеты.
Терминирующие кодоны узнаются специфическими белками — факторами терминации. Надежность распознавания этих триплетов невысока и поэтому терминирующий триплет иногда может прочитываться как кодон аминокислоты. Из-за этого терминирующие кодоны обычно дублируются. При этом первым обычно располагается кодон УАА, а уже за ним на близком расстоянии следует один из запасных терминирующих триплетов (УАГ или УГА).
Код в основном одинаков у всех животных организмов от вирусов до человека. В этом проявляется его универсальность, свидетельствующая о древности его происхождения и консервативности. Сходство кода у разных организмов является доказательством того, что все живые организмы в процессе эволюции произошли от единого предка.
Процессы синтеза ДНК, РНК и белков практически одинаковы у всех живых организмов. Поэтому в основе эволюции организмов были заложены механизмы образования новых генов, обусловливающих синтез различных функциональных белков, отличающихся по структуре и функциям. Это способствовало появлению разнообразных живых организмов на нашей планете.
248
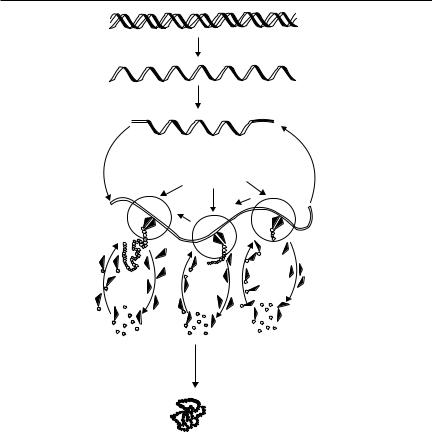
15.1. Биосинтез полипептидной цепочки белка на рибосоме
ДНК
транскрипция
преРНК
сплайсинг
мРНК
трансляция
рибосомы
тРНК
тРНК |
тРНК |
Рис. 15.1. Общая схема биосинтеза белка (Спирин, 1986)
Всего 20 разновидностей аминокислот могут быть включены в первичную структуру белка. Радикалы этих аминокислот проявляют гидрофобные, полярные незаряженные и заряженные (положительно и отрицательно) свойства. В полярной среде белки приобретают устойчивую структуру, при этом заряженные аминокислотные остатки располагаются на поверхности белковой глобулы. Тогда как полярные и гидрофобные аминокислотные остатки обращены преимущественно вовнутрь белка. Таким образом, сформированный белок приобретает функциональную активность. Причем поверхностные аминокислотные остатки несут информацию о клетке, реализация которой обеспечивает их упорядоченное в ней расположение и функционирование.
Функционально активным белок становится в процессе фолдинга, после завершения которого белки приобретают способность выполнять
249

Глава 15. Метаболизм белков
|
|
|
|
|
|
|
Та б л и ц а |
1 5 . 1 |
||
|
|
|
Генетический код |
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
У |
Ц |
|
А |
|
Г |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
У |
УУУ |
|
УЦУ |
|
УАУ |
УГУ |
|
|
У |
|
|
|
Фен |
|
|
Тир |
|
Цис |
|
|
|
|
УУЦ |
|
УЦЦ |
|
УАЦ |
УГЦ |
|
|
Ц |
|
|
|
|
|
Сер |
|
|
|
|
|
|
|
УУА |
|
УЦА |
|
УАА |
УГА |
Терм |
|
А |
|
|
|
Лей |
|
|
Терм |
|
|
|
|
|
|
УУГ |
|
УЦГ |
|
УАГ |
УГГ |
Три |
|
Г |
|
Ц |
ЦУУ |
|
ЦЦУ |
|
ЦАУ |
ЦГУ |
|
|
У |
|
|
|
|
|
|
Гис |
|
|
|
|
|
|
ЦУЦ |
|
ЦЦЦ |
|
ЦАЦ |
ЦГЦ |
|
|
Ц |
|
|
|
Лей |
|
Про |
|
|
Арг |
|
|
|
|
ЦУА |
|
ЦЦА |
|
ЦАА |
ЦГА |
|
|
А |
|
|
|
|
|
|
Глн |
|
|
|
|
|
|
ЦУГ |
|
ЦЦГ |
|
ЦАГ |
ЦГГ |
|
|
Г |
|
А |
АУУ |
|
АЦУ |
|
ААУ |
АГУ |
|
|
У |
|
|
|
|
|
|
Асн |
|
Сер |
|
|
|
|
АУЦ |
Иле |
АЦЦ |
|
ААЦ |
АГЦ |
|
|
Ц |
|
|
|
|
|
Тре |
|
|
|
|
|
|
|
АУА |
|
АЦА |
|
ААА |
АГА |
|
|
А |
|
|
|
|
|
|
Лиз |
|
Арг |
|
|
|
|
АУГ |
Мет |
АЦГ |
|
ААГ |
АГГ |
|
|
Г |
|
Г |
ГУУ |
|
ГЦУ |
|
ГАУ |
ГГУ |
|
|
У |
|
|
|
|
|
|
Асп |
|
|
|
|
|
|
ГУЦ |
|
ГЦЦ |
|
ГАЦ |
ГГЦ |
|
|
Ц |
|
|
|
Вал |
|
Ала |
|
|
Гли |
|
|
|
|
ГУА |
|
ГЦА |
|
ГАА |
ГГА |
|
|
А |
|
|
|
|
|
|
Глу |
|
|
|
|
|
|
ГУГ |
|
ГЦГ |
|
ГАГ |
ГГГ |
|
|
Г |
|
|
|
|
|
|
|
|
|
|
|
|
специализированные функции. При этом только небольшая группа аминокислот, расположенных на поверхности белковой глобулы, способна участвовать в функциональном действии белка.
Биосинтез белка представляет собой циклический энергозависимый многоступенчатый процесс, протекающий с участием мРНК, тРНК, рибосомы и различных инициирующих факторов. Вначале процесса происходит активирование аминокислот, а затем при наличии мРНК формирование функционально активной рибосомы. Процесс завершается образованием полипептидной цепочки, в составе которой количественный и качественный состав аминокислот будет определять индивидуальное строение будущего белка, его функциональные возможности и место локализации в структуре клеток.
250

15.1. Биосинтез полипептидной цепочки белка на рибосоме
Активирование аминокислот. Этот этап сопровождается присоединением аминокислоты, предназначенной для включения в состав полипептидной цепочки, к соответствующей тРНК. Процесс катализируют специализированные ферменты — аминоацил-тРНК-синтетазы. Ферменты обладают высокой специфичностью как к аминокислоте, так и к соответствующей тРНК.
Процесс ацилирования аминокислот протекает в цитоплазме клетки в две стадии. Сначала происходит образование аминоациладенилата. При этом карбоксильная группа аминокислоты связывается ангидридной связью с 5′-фосфатной группой АМФ с выделением пирофосфата.
|
|
АТФ |
ФФн |
||
R–CH–COOH |
|
|
R–CH–CO O-АМФ |
||
|
|
||||
|
|
|
|
|
|
NH2 |
|
NH2 |
|||
Аминокислота |
|
Аминоациладенилат |
|||
Затем происходит перенос аминоацильного остатка на специфическую тРНК с образованием сложноэфирной связи между карбоксильной группой аминокислоты и 3′-ОН-группой концевого аденозинового остатка тРНК.
R–CH–CO O-АМФ + тРНК |
|
R–CH–CO O-тРНК + АМФ |
||
|
||||
|
|
|
|
|
NH2 |
|
NH2 |
||
|
|
|
Аминоацил-тРНК |
|
Таким образом, результатом реакции этерификации является образование соответствующих аминоацил-тРНК, способных участвовать в процессе синтеза полипептидной цепочки.
Трансляция. Матричный синтез белка протекает с участием рибосомы. В результате этого процесса осуществляется перевод информации, заложенной в последовательности нуклеотидов мРНК, в последовательность аминокислотных остатков полипептидной цепи. Трансляция протекает в три этапа: инициация (начало процесса), элонгация (развитие процесса) и терминация (окончание процесса).
При наличии мРНК, которые поступают из ядра в цитоплазму через ядерные поры, происходит сборка рибосом. Началом синтеза белка является присутствие в составе мРНК концевых триплетов АУГ или ГУГ. Инициатором процесса служит метионил-тРНК, которая реагирует с факторами инициации elF-5, elF-2, elF-3, с 40S-субчастицей рибосомы и мРНК.
О
H3C–S–CH2–CH2–CH–C O-тРНК
NH2
Метионил-тРНК
251

Глава 15. Метаболизм белков
Процесс инициации протекает следующим образом. Инициирующий фактор elF-3 соединяется с малой субчастицей рибосомы (40S), а фактор elF-2 соединяется с ГТФ. Затем к комплексу elF-2-ГТФ присоединяется метионил-тРНК. В дальнейшем к комплексу elF-2-ГТФ присоединяется 40S-субчастица с elF-3. В результате образуется комплекс 40S-elF-3-ГТФ- elF-2-метионил-тРНК. К этому комплексу присоединяется мРНК своим 5′-концом при участии elF-5. Завершается процесс сборки рибосомы присоединением большой субчастицы рибосомы (60S), что сопровождается высвобождением факторов инициации, а также ГДФ и Н3РО4. После этого факторы инициации могут быть использованы для инициации синтеза новых рибосом.
Синтез пептидов в митохондриях и хлоропластах протекает с участием формилметионил-тРНК.
О
H3C–S–CH2–CH2–CH–C O-тРНК
NH
СНО Формилметионил-тРНК
Начало элонгации характеризуется тем, что в Р-участке рибосомы находится метионил-тРНК. При этом А-участок свободен и в нем находится следующий кодон мРНК. Поэтому вновь поступающая аминоацил-тРНК имеет антикодон, соответствующий кодону мРНК в А-участке. Для осуществления распознавания и связывания аминоацил-тРНК в А-участке требуется ГТФ и фактор элонгации (EF-1). Специфичность связывания очередной аминоацил-тРНК в А-участке осуществляется благодаря соответствию кодона мРНК антикодону тРНК. После этого происходит образование пептидной связи в пептидил-трансферазном центре большой субчастицы рибосомы, которая формируется в результате взаимодействия аминогруппы с вновь поступившей в рибосому аминокислоты с карбоксилом предыдущей аминокислоты. Реакция катализируется пептидилтрансферазой, входящей в состав большой субчастицы рибосомы. После образования пептидной связи осуществляется передвижение (транслокация) мРНК в рибосоме на один кодон. В этом действии участвует фактор инициации (EF-2) и расходуется энергия молекулы ГТФ.
В результате транслокации мРНК перемещается на один триплет, а ди- пептидил-тРНК поступает в Р-участок, вытесняя инициаторную мети- онил-тРНК. При этом в А-участок встает следующий кодон. Процесс повторяется при наличии следующей аминоацил-тРНК. Считывание информации с мРНК идет в направлении 5′→3′полинуклеотидной цепочки, а полипептидная цепь растет от N-конца к С-концу.
252