

-
Уравнение Гольдмана – Ходжкина – Хаксли.
Мембрана в состоянии покоя проницаема не только для ионов калия, но и в небольшой степени для ионов натрия и хлора. Мембранный потенциал клеток представляет собой результирующую электродвижущих сил, генерируемых всеми этими тремя каналами диффузии. Проникновение натрия из окружающей жидкости внутрь клетки по концентрационному градиенту приводит к некоторому уменьшению мембранного потенциала. Диффузия через мембрану ионов хлора, содержание которых в межклеточной жидкости большинства тканей выше, чем в клетках, вызывает некоторое увеличение мембранного потенциала. Следовательно, для более точного вычисления потенциала покоя необходимо учитывать не только диффузию ионов калия, но и диффузию натрия и хлора. Уравнение Нернста позволяет описывать мембранный потенциал только в первом приближении; для более точного определения потенциала применяют обобщенное уравнение Гольдмана-Ходжкина-Хаксли:
Е = (RT/ z F)ln[(Рк [K.]i + PNa [Na]i + PCl [Cl]e) / (Рк [K.]e + PNa [Na]e + PCl [Cl]i)
где Pk, PNa, PCl — коэффициенты проницаемости мембраны для ионов калия, натрия и хлора; [К], [Na], [Cl] — их активности внутри (i) и вне (е) клетки. Уравнение Гольдмана-Ходжкина-Хаксли позволяет определять мембранный потенциал не только в состоянии покоя, но и при возбуждении клетки. Для этого необходимо знание коэффициентов проницаемости для ионов. По данным Ходжкина и Катца (1949), для аксона кальмара в состоянии покоя отношение коэффициентов проницаемости Рк: Рnа : РCl= 1 : 0,04 : 0,45.
-
Потенциал действия. Генерация потенциала действия.
Все
клетки возбудимых тканей при действии
различных раздражителей достаточной
силы способны переходить в состояние
возбуждения. К возбудимым относятся
нервная, мышечная и железистая ткани,
хотя четкой границы между возбудимыми
и другими тканями провести нельзя.
Возбудимость—это
способность клеток к быстрому ответу
на раздражение, проявляющемуся через
совокупность физических, физико-химических
процессов и функциональных изменений.
Обязательным
признаком
возбуждения
является
изменение электрического состояния
клеточной мембраны.
Опыт показывает, что возбужденный
участок клетки становится электроотрицательным
по отношению к невозбужденному,
что указывает на перераспределение
ионов в возбужденном участке. При
возбуждении оно имеет временный характер,
и после окончания возбуждения вновь
восстанавливается исходный потенциал
покоя. Потенциалом
действия
называется
общее
изменение разности потенциалов между
клеткой и средой, происходящее при
пороговом и сверхпороговом возбуждении
клеток.
Потенциалы
действия
обеспечивают_проведение _возбуждения
по нервным волокнам и инициируют процессы
с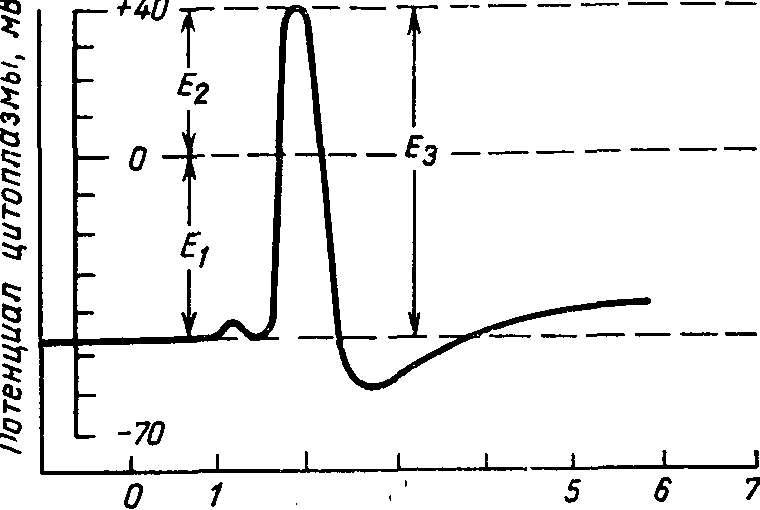 окращения
мышечных и секреции железистых клеток.
окращения
мышечных и секреции железистых клеток.
Время, мс
Рис. 1. Потенциал действия нервного волокна кальмара, зарегистрированный с помощью внутриклеточного электрода (по А. Ходжкину и А. Хаксли, 1965).
Е1 — потенциал покоя, E2 — мембранный потенциал при возбуждении, E3 - потенциал действия.
На рис. 1, где показано изменение мембранного потенциала при возбуждении, видно, что в состоянии покоя внутренняя сторона клеточной оболочки имеет отрицательный потенциал, примерно равный —45 мВ, тогда как при возбуждении мембранный потенциал становится равным +40 мВ. Таким образом, полное значение потенциала действия составляет 85 мВ, что значительно превышает величину потенциала покоя.
Возникновение потенциала действия связано с увеличением проницаемости мембраны для ионов натрия и последующим усилением диффузии этих ионов по концентрационному градиенту внутрь клетки, что приводит к изменению (уменьшению) мембранного потенциала. Независимо от того, чем это вызвано - уменьшение мембранного потенциала ниже критического уровня приводит к увеличению проницаемости мембраны для натрия, а увеличение проницаемости сопровождается усилением диффузии натрия в цитоплазму, что вызывает еще большую деполяризацию мембраны. Благодаря наличию положительной обратной связи деполяризация мембраны при возбуждении происходит с ускорением и поток ионов натрия в клетку все время возрастает.