

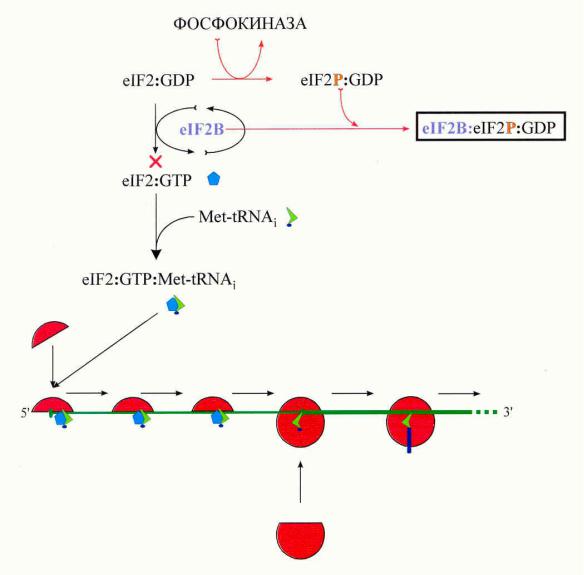
Цикл инициаторных факторов eIF2:GDP/GTP и eIF2B
|
e I F 2 : G T P : e I F 2 B |
M e t - t R N A i |
||||
G T P |
|
|
|
|
||
e I F 2 : e I F 2 B |
M e t - t R N A i |
: e I F 2 : G T P : e I F 2 B |
||||
G D P |
|
|
4 0 S S U B U N I T |
|||
|
|
|
|
|
|
|
|
|
e |
I F 2 B |
|
|
|
|
|
|
|
|
|
|
|
e I F 2 : G D P |
|
4 0 S : M e t - t R N A i : e I F 2 : G T P |
|
||
|
|
|||||
|
|
|
|
|||
8 0 S : M e t - t R N A i |
6 0 S S U B U N I T |
I N I T I A T I O N C O M P L E X
eIF2-киназы: фосфорилирование Ser51 -субъединицы eIF2
Индукторы eIF2-киназ:
(1)Недостаток Fe3+ и/или гема
(2)Вирусная инфекция
(3)Голодание
(4)Недостаток ростовых факторов
(5)Тепловой шок
Регуляция доступности
активного Met-tРНКi-связывающего фактора инициации eIF2 путем его фосфорилирования
eIF2P:GTP:eIF2B
e I F 2 : G T P : e I F 2 B |
M e t - t R N A i |
|
G T P |
|
|
eIF2P:eIF2B |
M e t - t R N A i : e I F 2 : G T P : e I F 2 B |
|
e I F 2 : e I F 2 B |
||
G D P |
|
4 0 S S U B U N I T |
|
e I F 2 B |
|
eIF2P:GDP |
4 0 S : M e t - t R N A i : e I F 2 : G T P |
|
e I F 2 : G D P |
||
P
8 0 S : M e t - t R N A i |
6 0 S S U B U N I T |
I N I T I A T I O N C O M P L E X
Активация гем-регулируемой eIF2-киназы (“HCR”):
Индуктор:
Недостаток Fe3+ и/или гема
|
|
HCR |
|
|
Inactive |
S |
Fe |
S |
|
dimer |
S |
S |
||
|
||||
|
|
HCR |
|
|
heme |
|
|
heme |
|
|
|
HCR |
|
SH SH
Active monomer
SH SH
HCR
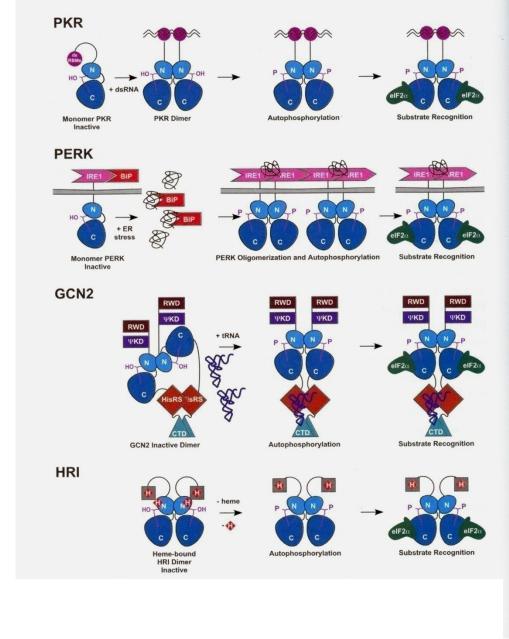
eIF2-киназы эукариот
Киназа, активируемая двуспиральной РНК
Киназа, активируемая при стрессе
(напр., при тепловом шоке)
Киназа, активируемая при голодании
Киназа, активируемая при недостатке гема и/или железа
Механизм тотального подавления инициации трансляции
eIF2-киназами

Вот теперь совсем Конец лекции
Регуляция доступности лимитирующего фактора инициации eIF4E/4F путем фосфорилирования/дефосфорилирования eIF4E-связывающего белка (4E-ВР)
[eIF4E – лимитирующий компонент аппарата инициации!]
|
+ eIF4A + eIF4G |
|
|
|
eIF4F |
|
eIF4E |
|
|
|
|||
|
|
|
||||
+ |
|
|
|
|
|
|
4E-BP |
|
eIF4E:4E-BP (inactive) |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
+ATP
фосфорилирование 4Е-ВР
Множественное фосфорилирование белка 4Е-ВР митоген-зависимой киназой (МАР), индуцированной инсулином или факторами роста,
приводит к диссоциации комплекса 4E-BP и освобождению активного eIF4Е.