

Строение и функции биологических мембран. Транспорт веществ через биологические мембраны
СТРОЕНИЕ И ФУНКЦИИ БИОЛОГИЧЕСКИХ МЕМБРАН. ЛИПИДЫ БИОМЕМБРАН. МЕХАНИЗМЫ ТРАНСПОРТА ВЕЩЕСТВ ЧЕРЕЗ БИОМЕМБРАНЫ
1. Многообразие мембранных структур и выполняемых ими функций.
2. Строение липидов, входящих в состав клеточных мембран: формулы фосфолипидов, гликолипидов, холестерола. Амфипатические свойства липидов мембран.
3. Белки мембран (интегральные, периферические): особенности структуры, свойства, функции. Взаимодействия белков и липидов в биологических мембранах.
4. Асимметрия мембран (примеры). Способность белков и липидов мембран к латеральной диффузии. Ограниченная возможность поперечной диффузии в мембранах.
5. Транспорт веществ через мембраны: простая и облегчённая диффузия, активный транспорт, экзо- и эндоцитоз, их особенности.
6. Na+, K+-АТФ-аза, механизм действия, роль в поддержании трансмембранного потенциала и возбудимости мембраны.
Ссылки по теме:
Березов Т.Т., Коровкин Б.Ф. Биологическая химия. Глава 9. Биомембраны и биоэнергетика. Основные принципы организации биомембран.
Раздел 9.1 |
Классификация липидов и их роль в организме. |
|
|
9.1.1. Из курса биоорганической химии известно, что липиды – это большая группа соединений, которые существенно различаются по своему химическому строению и биологической роли. Общими признаками липидов являются:
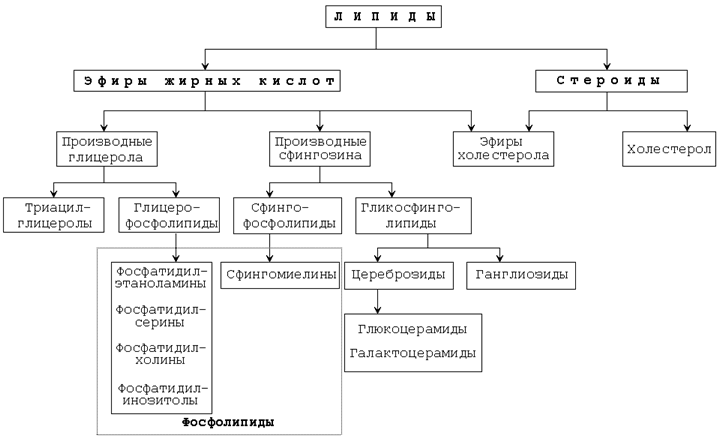
Основные классы липидов представлены на рисунке 9.1.
Рисунок 9.1. Основные классы липидов. 9.1.2. Липиды выполняют в организме ряд функций. Главными из них являются следующие:
|
|
Раздел 9.2 |
Классификация и функции биологических мембран. |
|
|
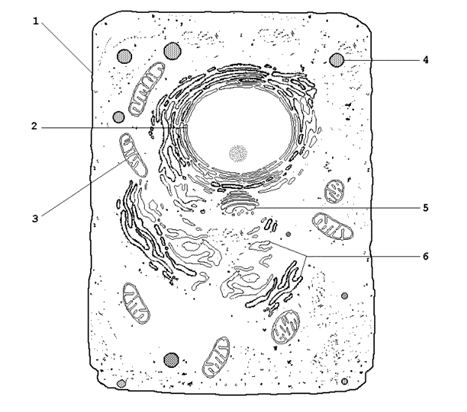
9.2.1. Биологические мембраны – сложные надмолекулярные структуры, окружающие все живые клетки и образующие в них замкнутые, специализированные компартменты – органеллы. Основные разновидности биологических мембран представлены на рисунке 9.2. Мембрану, ограничивающую цитоплазму клетки снаружи, называют цитоплазматической или плазматической мембраной (1). Название внутриклеточных мембран обычно происходит от названия ограничиваемых или образуемых ими субклеточных структур. Различают ядерную (2), митохондриальную (3), лизосомальную (4) мембраны, мембраны комплекса Гольджи (5), эндоплазматический ретикулум (6) и другие.
Рисунок 9.2. Схематическое изображение органелл животной клетки (объяснения в тексте). 9.2.2. Некоторые примеры функций биологических мембран. Плазматическая мембрана – ограничивает содержимое клетки от внешней среды; осуществляет контакт с другими клетками, получение, обработку и передачу информации внутрь клетки, поддержание постоянства внутренней среды. Ядерные мембраны (внешняя и внутренняя) – образуют ядерную оболочку, которая отделяет хромосомный материал от цитоплазматических органелл; через поры ядерной оболочки происходит транспорт белков и нуклеиновых кислот в ядро и из ядра. Митохондриальные мембраны – осуществляют преобразование энергии в ходе окислительного фосфорилирования, синтез АТФ. Лизосомальные мембраны – ограничивают гидролитические ферменты от цитоплазмы клетки, препятствуют самоперевариванию (аутолизу) клеток, способствуют поддержанию постоянства рН среды в лизосомах. Мембраны эндоплазматического ретикулума – принимают участие в образовании новых мембран, осуществляют синтез белков, липидов, полисахаридов, окисление гидрофобных метаболитов и ксенобиотиков. |
|
Раздел 9.3 |
Состав и строение мембран. |
||||||||
|
|||||||||
9.3.1. Все мембраны по своей организации и составу обнаруживают ряд общих свойств. Они:
9.3.2. Схема строения биологической мембраны, представлена на рисунке 9.3. Основу мембраны составляет липидный бислой – двойной слой молекул липидов, которые обладают свойствомамфифильности (содержат как гидрофильные, так и гидрофобные функциональные группы). В липидном бислое гидрофобные участки молекул взаимодействуют между собой, а гидрофильные участки обращены в окружающую водную среду.
Рисунок 9.3. Схема строения биологической мембраны (по Сингеру и Николсону). Мембранные липиды выполняют роль растворителя мембранных белков, создавая жидкую среду, в которой они могут функционировать. По степени влияния на структуру бислоя и по силе взаимодействия с ним мембранные белки делят на интегральные и периферические. Важнейшие особенности интегральных и периферических белков представлены в таблице 9.1.
Таблица 9.1 Характеристика мембранных белков
По выполняемым функциям белки в составе мембран делятся на
Количество белков в мембранах могут существенно отличаться. Например, в миелиновой мембране, предназначенной для изоляции нервных волокон, белки составляют только 25% массы мембраны, а в мембранах митохондрий, связанных с процессами окислительного фосфорилирования, на долю белков приходится около 75% массы. В плазматической мембране доля белков и липидов примерно одинаковы. Углеводы в составе мембран не представлены самостоятельными соединениями, а обнаруживаются только в соединении с белками (гликопротеины) или липидами (гликолипиды). Длина углеводных цепей колеблется от двух до восемнадцати остатков моносахаридов. Большая часть углеводов расположена на наружной поверхности плазматической мембраны. Функции углеводов в биомембранах – контроль за межклеточными взаимодействиями, поддержание иммунного статуса, рецепция, обеспечение стабильности белковых молекул в мембране. |
|||||||||

Раздел 9.4 |
Липидный состав мембран |
|
|
9.4.1. Как уже упоминалось (9.1), компонентами липидов являются остатки жирных кислот и одно- или многоатомных спиртов. Примеры жирных кислот, встречающихся в составе липидов мембран, представлены на рисунке 9.4. Выучите эти формулы.
Рисунок 9.4. Наиболее часто встречающиеся природные жирные кислоты. Основные особенности строения жирных кислот, входящих в состав природных жиров:
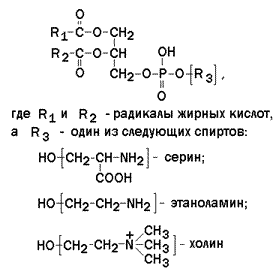
9.4.2. Большинство липидов в мембранах млекопитающих представлены фосфолипидами, гликосфинголипидами и холестеролом. Фосфолипиды в составе мембран подразделяются на две группы: глицерофосфолипиды и сфингомиелины. Глицерофосфолипиды – представляют собой сложные эфиры трёхатомного спирта глицерола, двух остатков жирных кислот и фосфорилированного аминоспирта. Общая формула глицерофосфолипида представлена на рисунке 9.5. Наиболее распространённым глицерофосфолипидом мембран является фосфатидилхолин:
В глицерофосфолипидах у второго углеродного атома глицерола обязательно находится остаток ненасыщенной жирной кислоты (в данном случае линолевой).
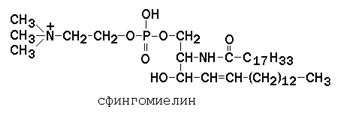
Рисунок 9.5. Общая формула глицерофосфолипидов. Сфингофосфолипиды (сфингомиелины) являются производными аминоспирта сфингозина (рисунок 9.6). Соединение сфингозина и жирной кислоты получило название церамид.
Рисунок 9.6. Структурные формулы сфингозина и его производных. В сфингомиелинах водород гидроксильной группы у первого углеродного атома в церамиде замещён на фосфохолин. Пример сфингомиелина, содержащего остаток олеиновой кислоты:
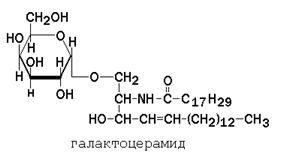
Гликолипиды также являются производными церамида, содержащими один или несколько остатков моносахаридов. Например, цереброзиды содержат в первом положении остаток глюкозы или галактозы:
а ганглиозиды содержат цепочку из нескольких остатков сахаров, одним из которых обязательно является сиаловая кислота. Холестерол (рисунок 9.7) – одноатомный циклический спирт. Это один из главных компонентов плазматической мембраны клеток млекопитающих, в меньшем количестве может присутствовать также в митохондриях, мембранах комплекса Гольджи, ядерных мембранах. Особенно много его в нервной ткани.
Рисунок 9.7. Структурные формулы холестерола и его эфира. 9.4.3. Как уже было сказано, характерной особенностью мембранных липидов является их амфифильность – наличие в молекуле одновременно гидрофобных и гидрофильных участков. Гидрофобная часть молекулы представлена остатками жирных кислот и боковой цепью сфингозина. Гидрофильные участки представлены в фосфолипидах фосфорилированным спиртом, а в гликолипидах – остатками сахаров. Амфифильность холестерола выражена слабо – циклическая структура и боковой радикал гидрофобны, и только гидроксильная группа гидрофильна. Амфифильность мембранных липидов определяет характер их поведения в водной среде. Слипание гидрофобных участков молекул приводит к образованию упорядоченных замкнутых структур – мицелл, в которых гидрофобные области защищены от воды, а гидрофильные обращены в водную среду. Молекулы холестерола встраиваются между радикалами жирных кислот гидрофобной части бислоя, а его гидроксильная группа примыкает к гидрофильным головкам фосфолипидов. Такая структура, стабилизированная нековалентными гидрофобными взаимодействиями, термодинамически очень устойчива и лежит в основе формирования биологических мембран. 9.4.4. Замкнутый липидный бислой определяет основные свойства мембран: 1) текучесть – зависит от соотношения насыщенных и ненасыщенных жирных кислот в составе мембранных липидов. Гидрофобные цепочки насыщенных жирных кислот ориентированы параллельно друг другу и образуют жёсткую кристаллическую структуру (рисунок 9.8, а). Ненасыщенные жирные кислоты, имеющие изогнутую углеводородную цепь, нарушают компактность упаковки и придают мембране бóльшую жидкостность (рисунок 9.8, б). Холестерол, встраиваясь между жирными кислотами, уплотняет их и повышает жёсткость мембран.
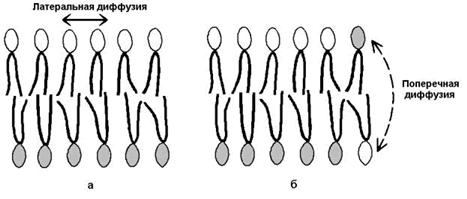
Рисунок 9.8. Влияние жирнокислотного состава фосфолипидов на текучесть липидного бислоя. 2) латеральная диффузия – свободное перемещение молекул относительно друг друга в плоскости мембран (рисунок 9.9,а).
Рисунок 9.9. Виды перемещений фосфолипидных молекул в липидном бислое. 3) ограниченная способность к поперечной диффузии (переходу молекул из наружного слоя во внутренний и наоборот, см. рисунок 9.9, б), что способствует сохранению асимметрии – структурно-функциональных различий наружного и внутреннего слоёв мембраны. 4) непроницаемость замкнутого бислоя для большинства водорастворимых молекул. |
|





Раздел 9.5 |
Трансмембранный перенос веществ. |
|
|
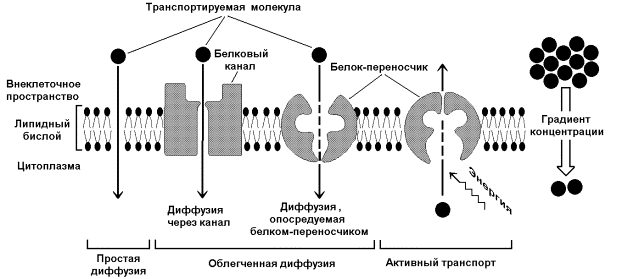
9.5.1. Одна из главных функций мембран – участие в переносе веществ. Этот процесс обеспечивается при помощи трёх основных механизмов: простой диффузией, облегчённой диффузией и активным транспортом (рисунок 9.10). Запомните важнейшие особенности этих механизмов и примеры транспортируемых веществ в каждом случае.
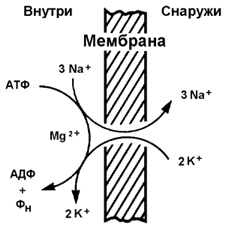
Рисунок 9.10. Механизмы транспорта молекул через мембрану Простая диффузия - перенос веществ через мембрану без участия специальных механизмов. Транспорт происходит по градиенту концентрации без затраты энергии. Путём простой диффузии транспортируются малые биомолекулы – Н2О, СО2, О2, мочевина, гидрофобные низкомолекулярные вещества. Скорость простой диффузии пропорциональна градиенту концентрации. Облегчённая диффузия - перенос веществ через мембрану при помощи белковых каналов или специальных белков-переносчиков. Осуществляется по градиенту концентрации без затраты энергии. Транспортируются моносахариды, аминокислоты, нуклеотиды, глицерол, некоторые ионы. Характерна кинетика насыщения – при определённой (насыщающей) концентрации переносимого вещества в переносе принимают участие все молекулы переносчика и скорость транспорта достигает предельной величины. Активный транспорт – также требует участия специальных белков-переносчиков, но перенос происходит против градиента концентрации и поэтому требует затраты энергии. При помощи этого механизма через клеточную мембрану транспортируются ионы Na+, K+, Ca2+, Mg2+, через митохондриальную – протоны. Для активного транспорта веществ характерна кинетика насыщения. 9.5.2. Примером транспортной системы, осуществляющей активный транспорт ионов, является Na+,K+-аденозинтрифосфатаза (Na+,K+-АТФаза или Na+,K+-насос). Этот белок находится в толще плазматической мембраны и способен катализировать реакцию гидролиза АТФ. Энергия, выделяемая при гидролизе 1 молекулы АТФ, используется для переноса 3 ионов Na+ из клетки во внеклеточное пространство и 2 ионов К+ в обратном направлении (рисунок 9.11). В результате действия Na+,K+-АТФазы создаётся разность концентраций между цитозолем клетки и внеклеточной жидкостью. Поскольку перенос ионов неэквивалентен, то возникает разность электрических потенциалов. Таким образом, возникает электрохимический потенциал, который складывается из энергии разности электрических потенциалов Δφ и энергии разности концентраций веществ ΔС по обе стороны мембраны.
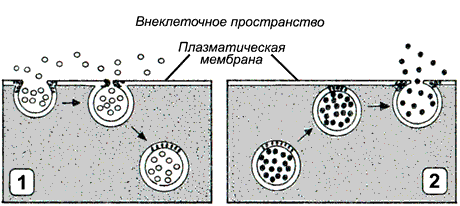
Рисунок 9.11. Схема Na+, K+-насоса. 9.5.3. Перенос через мембраны частиц и высокомолекулярных соединений Наряду с транспортом органических веществ и ионов, осуществляемым переносчиками, в клетке существует совершенно особый механизм, предназначенный для поглощения клеткой и выведения из неё высокомолекулярных соединений при помощи изменения формы биомембраны. Такой механизм называют везикулярным транспортом.
Рисунок 9.12. Типы везикулярного транспорта: 1 - эндоцитоз; 2 - экзоцитоз. При переносе макромолекул происходит последовательное образование и слияние окружённых мембраной пузырьков (везикул). По направлению транспорта и характеру переносимых веществ различают следующие типы везикулярного транспорта: Эндоцитоз (рисунок 9.12, 1) — перенос веществ в клетку. В зависимости от размера образующихся везикул различают: а) пиноцитоз — поглощение жидкости и растворённых макромолекул (белков, полисахаридов, нуклеиновых кислот) с помощью небольших пузырьков (150 нм в диаметре); б) фагоцитоз — поглощение крупных частиц, таких, как микроорганизмы или обломки клеток. В этом случае образуются крупные пузырьки, называемые фагосомами диаметром более 250 нм. Пиноцитоз характерен для большинства эукариотических клеток, в то время как крупные частицы поглощаются специализированными клетками - лейкоцитами и макрофагами. На первой стадии эндоцитоза вещества или частицы адсорбируются на поверхности мембраны, этот процесс происходит без затраты энергии. На следующей стадии мембрана с адсорбированным веществом углубляется в цитоплазму; образовавшиеся локальные впячивания плазматической мембраны отшнуровываются от поверхности клетки, образуя пузырьки, которые затем мигрируют внутрь клетки. Этот процесс связан системой микрофиламентов и является энергозависимым. Поступившие в клетку пузырьки и фагосомы могут сливаться с лизосомами. Содержащиеся в лизосомах ферменты расщепляют вещества, содержащиеся в пузырьках и фагосомах до низкомолекулярных продуктов (аминокислот, моносахаридов, нуклеотидов), которые транспортируются в цитозоль, где они могут быть использованы клеткой. Экзоцитоз (рисунок 9.12, 2) — перенос частиц и крупных соединений из клетки. Этот процесс, как и эндоцитоз, протекает с поглощением энергии. Основными разновидностями экзоцитоза являются: а) секреция - выведение из клетки водорастворимых соединений, которые используются или воздействуют на другие клетки организма. Может осуществляться как неспециализированными клетками, так и клетками эндокринных желёз, слизистой желудочно-кишечного тракта, приспособленными для секреции производимых ими веществ (гормонов, нейромедиаторов, проферментов) в зависимости от определённых потребностей организма. Секретируемые белки синтезируются на рибосомах, связанных с мембранами шероховатого эндоплазматического ретикулума. Затем эти белки транспортируются к аппарату Гольджи, где они модифицируются, концентрируются, сортируются, и затем упаковываются в пузырьки, которые отщепляются в цитозоль и в дальнейшем сливаются с плазматической мембраной, так что содержимое пузырьков оказывается вне клетки. В отличие от макромолекул, секретируемые частицы малых размеров, например, протоны, транспортируются из клетки при помощи механизмов облегчённой диффузии и активного транспорта. б) экскреция - удаление из клетки веществ, которые не могут быть использованы (например, удаление в ходе эритропоэза из ретикулоцитов сетчатой субстанции, представляющей собой агрегированные остатки органелл). Механизм экскреции, по-видимому, состоит в том, что вначале выделяемые частицы оказываются в цитоплазматическом пузырьке, который затем сливается с плазматической мембраной. |
|
http://dendrit.ru/page/show/mnemonick/stroenie-i-funkcii-biologicheskih-membra
Биологические мембраны, их свойства и функции
Биологические мембраны, их свойства и функции
Строение биологических мембран. Одной из основных особенностей всех эукариотических клеток является изобилие и сложность строения внутренних мембран. Мембраны отграничивают цитоплазму от окружающей среды, а также формируют оболочки ядер, митохондрий и пластид. Они образуют лабиринт эндр-плазматического ретикулума и уплощенных пузырьков в виде стопки, составляющих комплекс Гольджи. Мембраны образуют лизосомы, крупные и мелкие вакуоли растительных и грибных клеток, пульсирующие вакуоли простейших. Все эти структуры представляют собой компартменты (отсеки), предназначенные для тех или иных специализированных процессов и циклов. Следовательно, без мембран существование клетки невозможно.
Плазматическая мембрана, или плазмалемма, — наиболее постоянная, основная, универсальная для всех клеток мембрана. Она представляет собой тончайшую (около 10 нм) пленку, покрывающую всю клетку. Плазмалемма состоит из молекул белков и фосфолипидов (рис. 1.6). Молекулы фосфолипидов расположены в два ряда — гидрофобными концами внутрь, гидрофильными головками к внутренней и внешней водной среде. В отдельных местах бислой (двойной слой) фосфолипидов насквозь пронизан белковыми молекулами (интегральные белки). Внутри таких белковых молекул имеются каналы — поры, через которые проходят водорастворимые вещества. Другие белковые молекулы пронизывают бислой липидов наполовину с одной или с другой стороны (полуинтегральные белки). На поверхности мембран эукариотических клеток имеются периферические белки. Молекулы липидов и белков удерживаются благодаря гидрофильно-гидрофобным взаимодействиям.

Рис. 1.6. Схема строения мембраны: а — трехмерная модель; б — плоскостное изображение; 1 — белки, примыкающие к липидному слою (А), погруженные в него (Б) или пронизывающие его насквозь (В); 2 — слои молекул липидов; 3 —гликопротеины; 4 — гликолипиды; 5 — гидрофильный канал, функционирующий как пора.
В состав плазматической мембраны эукариотических клеток входят также полисахариды. Их короткие, сильно развлетвленные молекулы ковалентно связаны с белками, образуя гликопротеины, или с липидами (гликолипиды). Содержание полисахаридов в мембранах составляет 2-—10% по массе. Полисахаридный слой толщиной 10—20 нм, покрывающий сверху плазмалемму животных клеток, получил название гликокаликс.
Свойства и функции мембран.
Все клеточные мембраны представляют собой подвижные текучие структуры, поскольку молекулы липидов и белков не связаны между собой ковалентными связями и способны достаточно быстро перемещаться в плоскости мембраны. Благодаря этому мембраны могут изменять свою конфигурацию, т. е. обладают текучестью.
Мембраны — структуры очень динамичные. Они быстро восстанавливаются после повреждения, а также растягиваются и сжимаются при клеточных движениях.
Мембраны разных типов клеток существенно различаются как по химическому составу, так и по относительному содержанию в них белков, гликопротеинов, липидов, а следовательно, и по характеру имеющихся в них рецепторов. Каждый тип клеток поэтому характеризуется индивидуальностью, которая определяется в основном гликопротеинами. Разветвленные цепи гликопротеинов, выступающие из клеточной мембраны, участвуют в распознава-нии факторов внешней среды, а также во взаимном узнавании родственных клеток. Например, яйцеклетка и сперматозоид узнают друг друга по гликопротеинам клеточной поверхности, которые подходят другкдругу как отдельные элементы цельной структуры. Такое взаимное узнавание — необходимый этап, предшествующий оплодотворению.
Подобное явление наблюдается в процессе дифференциров-ки тканей. В этом случае сходные по строению клетки с помощью распознающих участков плазмалеммы правильно ориентируются относительно друг друга, обеспечивая тем самым их сцепление и образование тканей. С распознаванием связана и регуляция транспорта молекул и ионов через мембрану, а также иммунологический ответ, в котором гликопротеины играют роль антигенов. Сахара, таким образом, могут функционировать как информационные молекулы (подобно белкам и нуклеиновым кислотам). В мембранах содержатся также специфические рецепторы, переносчики электронов, преобразователи энергии, ферментные белки. Белки участвуют в обеспечении транспорта определенных молекул внутрь клетки или из нее, осуществляют структурную связь цитоскелета с клеточными мембранами или же служат в качестве рецепторов для получения и преобразования химических сигналов из окружающей среды. Важнейшим свойством мембраны является также избирательная проницаемость. Это значит, что молекулы и ионы проходят через нее с различной скоростью, и чем больше размер молекул, тем меньше скорость прохождения их через мембрану. Это свойство определяет плазматическую мембрану как осмотический барьер. Максимальной проникающей способностью обладает вода и растворенные в ней газы; значительно медленнее проходят сквозь мембрану ионы. Диффузия воды через мембрану называется осмосом.
Существует несколько механизмов транспорта веществ через мембрану.
Диффузия —проникновение веществ через мембрану по градиенту концентрации {из области, где их концентрация выше, в область, где их концентрация ниже). Диффузный транспорт веществ (воды, ионов) осуществляется при участии белков мембраны, в которых имеются молекулярные поры, либо при участии липидной фазы (для жирорастворимых веществ).
При облегченной диффузии специальные мембранные белки-переносчики избирательно связываются с тем или иным ионом или молекулой и переносят их через мембрану по градиенту концентрации.
Активный транспорт сопряжен с затратами энергии и служит для переноса веществ против их градиента концентрации. Оносуществляется специальными белками-переносчиками, образующими так называемые ионные насосы. Наиболее изученным является Na-/ К--насос в клетках животных, активно выкачивающих ионы Na+ наружу, поглощая при этом ионы К-. Благодаря этому в клетке поддерживается большая концентрация К- и меньшая Na+ по сравнению с окружающей средой. На этот процесс затрачивается энергия АТФ.
В результате активного транспорта с помощью мембранного насоса в клетке происходит также регуляция концентрации Mg2-и Са2+. В процессе активного транспорта ионов в клетку через цито-плазматическую мембрану проникают различные сахара, нукле-отиды, аминокислоты. Макромолекулы белков, нуклеиновых кислот, полисахаридов, липопротеидные комплексы и др. сквозь клеточные мембраны не проходят, в отличие от ионов и мономеров. Транспорт макромолекул, их комплексов и частиц внутрь клетки происходит совершенно иным путем — посредством эндоцитоза. При эндоци-тозе {эндо... — внутрь) определенный участок плазмалеммы захватывает и как бы обволакивает внеклеточный материал, заключая его в мембранную вакуоль, возникшую вследствие впя-чивания мембраны. В дальнейшем такая вакуоль соединяется с лизосомой, ферменты которой расщепляют макромолекулы до мономеров. Процесс, обратный эндоцитозу, — экзоцитоз (экзо... — наружу). Благодаря ему клетка выводит внутриклеточные продукты или непереваренные остатки, заключенные в вакуоли или пузырьки. Пузырек подходит к цитоплазматической мембране, сливается с ней, а его содержимое выделяется в окружающую среду. Гак выводятся пищеварительные ферменты, гормоны, гемицел-люлоза и др.
Таким образом, биологические мембраны как основные структурные элементы клетки служат не просто физическими границами, а представляют собой динамичные функциональные поверхности. На мембранах органелл осуществляются многочисленные биохимические процессы, такие как активное поглощение веществ, преобразование энергии, синтез АТФ и др.
Функции биологических мембран следующие:
Отграничивают содержимое клетки от внешней среды и содержимое органелл от цитоплазмы.
Обеспечивают транспорт веществ в клетку и из нее, из цитоплазмы в органеллы и наоборот.
Выполняют роль рецепторов (получение и преобразование сит-налов из окружающей среды, узнавание веществ клеток и т. д.).
Являются катализаторами (обеспечение примембранных химических процессов).
Участвуют в преобразовании энергии.
Источник : Н.А. Лемеза Л.В.Камлюк Н.Д. Лисов "Пособие по биологии для поступающих в ВУЗы"
http://uclg.ru/education/biologiya/10_klass/kletka_-_edinitsa_jivogo/struktura_i_funktsii_kletki/lecture_lec_biologicheskie_membranyi__ih_svoystva_i_funktsii.html
1.Биология как наука о закономерностях и механизмах жизнедеятельности и развития организмов , её задачи. Объект и методы исследования
Термин биология (от греч. биос — жизнь, логос — наука) введен в начале XIX в. независимо Ж.-Б. Ламарком и Г. Тревиранусом для обозначения науки о жизни как особом явлении природы. В настоящее время его используют и в ином смысле, относя к группам организмов, вплоть до вида (биология микроорганизмов, биология северного оленя, биология человека), биоценозам (биология арктического бассейна), отдельным структурам (биология клетки). Предметом биологии как учебной дисциплины служит жизнь во всех ее проявлениях: строение, физиология, поведение, индивидуальное (онтогенез) и историческое (эволюция, филогенез) развитие организмов, их взаимоотношение друг с другом и с окружающей средой. Основными задачами биологии являются: 1) раскрытие сущности жизни и ее проявления с целью познания ее; 2) изучение живых существ и закономерности их в живой природе.Основные методы исследования в биологии являются:
Эмпирические – наблюдение, эксперимент.
Теоретические – сравнительный, исторический, моделирования, статическая обработка, формулировка гипотез, законов и теорий и др.
Наблюдение дает возможность описать биологические объекты и явления. При этом используются инструментальные методы (микроскопия, электрография, рентгенография и др)
Эксперимент – исследователи искусственно создают ситуации, которые помогают изучать свойства биологических объектов. Также используются инструментальные методы: электрофорез, хроматография, культивирование тканей и др.
Сравнение и обобщение – позволяют найти общие закономерности для нескольких явлений. С помощью этого метода была основана систематика, создана клеточная теория.
Исторический метод – позволяет на основе данных о современном органическом мире и его прошлом познать процессы развития природы.
Моделирование – имитирует отдельные биологические процессы. Например аквариум – модель природной экологической системы.
2. Дайте определение жизни. Охарактеризуйте свойства живого. Назовите формы жизни.
Жизнь – это макромолекулярная открытая система, которой свойственна иерархическая организация, способность к самовозобновлению, обмен веществ и тонко регуляторный процесс.
Свойства живого:
1. Самовозобновление, которое связано с постоянным обменом вещества и энергии, и в основе которого лежит способность хранить и использовать биологическую информацию в виде уникальных информационных молекул: белков и нуклеиновых кислот.
2. Самовоспроизведение, которое обеспечивает преемственность между поколениями биологических систем
3. Саморегуляция, которая основана на потоке вещества, энергии и информации
4. Большинство химических процессов в организме находятся не в динамичном состоянии
5. Живые организмы способны к росту .
3. Эволюционно-обусловленные уровни организации биологических систем.
1.Молекулярно – генетический.
На этом уровне изучаются физико – химические процессы, происходящие в организме – синтез и разложение белков, липидов, нуклеиновых кислот, обмен веществ и энергии, копирование генетической информации. Элементарной единицей на молекулярно-генетическом уровне служит ген, в котором записан определенный объем биологической наследственной информации. Элементарное явление на этом уровне – редупликация (самовоспроизведение) ДНК, в процессе которой могут возникать нарушения, изменяющие смысл генетической информации, приводящие к изменчивости. Биологическая информация, заключающаяся в молекулах ДНК, не участвует непосредственно в процессах жизнедеятельности. Она переходит в действующую форму, будучи перенесена в молекулы белков. Отмеченный перенос осуществляется благодаря механизму матричного синтеза, в котором исходная ДНК служит, как и в случае с редупликацией, матрицей (формой), но для образования не дочерней молекулы ДНК, а матричной РНК, контролирующей биосинтез белков. В основе этого процесса лежит принцип комплементарности. Это дает основание причислить матричный синтез информационных макромолекул также к элементарному явлению на молекулярно-генетическом уровне организации жизни.
2.Клеточный.
Элементарная структурная функциональная единица – клетка. Элементарное явление представлено реакциями клеточного метаболизма, составляющими основу потоков энергии, веществ и информации. Благодаря деятельности клетки поступающие извне вещества превращаются в субстраты и энергию, которые используются (в соответствии с имеющейся генетической информацией) в процессе биосинтеза белков и других соединений, необходимых организму. Таким образом, на клеточном уровне сопрягаются механизмы передачи биологической информации и превращения веществ и энергии. Элементарное явление на этом уровне служит энергетической и вещественной основой жизни на всех других уровнях ее организации.
3. Организменный.
Элементарной единицей организменного уровня является особь( организм) в ее развитии от момента зарождения до прекращения существования в качестве живой системы, что позволяет также назвать этот уровень онтогенетическим.
Закономерность изменения организма в индивидуальном развитии составляют элементарное явление данного уровня.
4. Популяционно – видовой.
Элементарной единицей популяционно-видового уровня служит популяция — совокупность особей одного вида. Объединение особей в популяцию происходит благодаря общности генофонда, используемого в процессе полового размножения для создания генотипов особей следующего поколения. Популяция в силу возможности межпопуляционных скрещиваний представляет собой открытую генетическую систему. Действие на генофонд популяции элементарных эволюционных факторов, таких, как мутационный процесс, колебания численности особей, естественный отбор, приводит к эволюционно значимым изменениям генофонда, которые представляют элементарные явления на данном уровне.
5. Биогеоценотический и биосферный.
В процессе совместного исторического развития на определенной территории организмов разных систематических групп образуются динамичные, устойчивые во времени сообщества — биогеоценозы, которые служат элементарной единицей биогеоценотического (экосистемного) уровня. Элементарное явление на рассматриваемом уровне представлено потоками энергии и круговоротами веществ. Ведущая роль в этих круговоротах и потоках принадлежит живым организмам.
Биогеоценоз — это открытая в вещественном и энергетическом плане система. Биогеоценозы, различаясь по видовому составу и характеристикам абиотической своей части, объединены на планете в единый комплекс — область распространения жизни, или биосферу. Биосфера – это совокупность всех биогеоценозов, образующих единый комплекс, охватывающий все явления жизни на планете.
https://studfiles.net/preview/5811033/