

вся инфа по нервам истории лекции рефераты / Топический диагноз в неврологии
.PDF248 5 Промежуточный мозг
пример, при грудной жабе, боль часто проецируется в соответствующие дер5 матомы.
Эти кожные зоны Геда имеют важ5 ное диагностическое значение (см. рис. 5.31). Импульсы, идущие от кожи, ока5 зывают влияние на 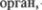 имеющий соответствующую висцеральную иннер5 вацию, и наоборот. Ясно, что в спинном мозге на уровне релейных нейронов должны существовать определенные
имеющий соответствующую висцеральную иннер5 вацию, и наоборот. Ясно, что в спинном мозге на уровне релейных нейронов должны существовать определенные
связи между афферентными соматичес5 кими волокнами и висцеральными реф5 лекторными дугами; в противном слу5 чае трудно 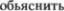 терапевтический эффект горячих
терапевтический эффект горячих 
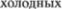
 аппликаций, различного рода оберты5 ваний, мазей, образования волдырей и других аналогичных лечебных меро5 приятий. Довольно часто они уменьша5 ют боль в органе, имеющем автоном5 ную (вегетативную) иннервацию.
аппликаций, различного рода оберты5 ваний, мазей, образования волдырей и других аналогичных лечебных меро5 приятий. Довольно часто они уменьша5 ют боль в органе, имеющем автоном5 ную (вегетативную) иннервацию.

6 Базальные ядра и экстрапирамидная система
Базальные ядра
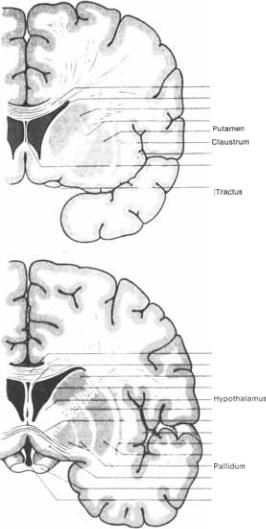
Термин базальные ядра (nuclei basales) относится к скоплениям серого веще5 ства в толще полушарий большого моз5 га. Они происходят из теленцефаличе5 ского ганглионарного бугорка эмбрио5 на. Эти ядра состоят из хвостатого ядра (nucleus caudatus), скорлупы (putamen),
ограды (claustrum) иминдалевидного те$ ла (corpus amygdaloideum). Последняя структура уже описана в разделе по лим5 бической системе.
Хвостатое ядро и скорлупа хотя и разделены внутренней капсулой (capsula interna), представляют собой одно ядро и имеют одинаковую гистологическую структуру (плотно расположенные мел5 кие нейроны и отдельные большие мультиполярные 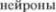 между ни5
между ни5
 Разделение хвостатого ядра и скор5 лупы неполное — они связаны между собой мостиками серого вещества, про5 бивающимися через волокна внутрен5 ней капсулы и содержащими два вы5 шеупомянутых вида нейронов (рис. 6.1, 6.2, 6.3).
Разделение хвостатого ядра и скор5 лупы неполное — они связаны между собой мостиками серого вещества, про5 бивающимися через волокна внутрен5 ней капсулы и содержащими два вы5 шеупомянутых вида нейронов (рис. 6.1, 6.2, 6.3).  где волокна внутренней капсулы иссякают, ядра со5 единены между собой довольно солид5 ным скоплением нервных клеток, на5 зываемым дном полосатого тела.
где волокна внутренней капсулы иссякают, ядра со5 единены между собой довольно солид5 ным скоплением нервных клеток, на5 зываемым дном полосатого тела.
Термин полосатое тело или
атум (striatum) обычно используется для обозначения обоих ядер. Полосами, о которых идет речь в их названии, являются множественные белого цвета
скопления миелинизированных воло5 кон, контрастирующих на фоне бледно5 го (вследствие слабой миелинизации) серого вещества и конвергирующих в направлении  шара.
шара.  ядро и скорлупа составляют неостриа$ тум (neostriatum), филогенетически бо5 лее более старый бледный шар называ5 ют палеостриатум
ядро и скорлупа составляют неостриа$ тум (neostriatum), филогенетически бо5 лее более старый бледный шар называ5 ют палеостриатум 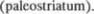 Час5 то о бледном шаре и скорлупе говорят как об одном образовании, называемом
Час5 то о бледном шаре и скорлупе говорят как об одном образовании, называемом
чечевицеобразным ядром (nucleus
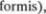 которое клином вдается между наружной и внутренней капсулой (см. рис. 6.4). Употребляя этот термин сле5 дует иметь ввиду, что два
которое клином вдается между наружной и внутренней капсулой (см. рис. 6.4). Употребляя этот термин сле5 дует иметь ввиду, что два  этого ядра имеют различное происхож5 дение, структуру и функцию. Скорлупа, как и хвостатое ядро, происходит из матрикса вокруг боковых желудочков и относится к неокортексу. Бледный шар (globus pallidus) является частью диэн5 цефалона, происходит из матрикса вок5 руг третьего желудочка и имеет общее происхождение с субталамическим яд5 ром (Richter, 1965).
этого ядра имеют различное происхож5 дение, структуру и функцию. Скорлупа, как и хвостатое ядро, происходит из матрикса вокруг боковых желудочков и относится к неокортексу. Бледный шар (globus pallidus) является частью диэн5 цефалона, происходит из матрикса вок5 руг третьего желудочка и имеет общее происхождение с субталамическим яд5 ром (Richter, 1965).
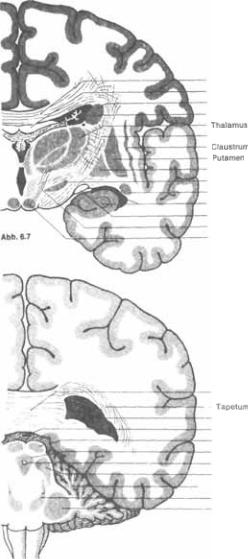
Хвостатое ядро повторяет очерта5 ния бокового желудочка и имеет форму открытого эллипса (см. рис. 6.2 и 6.3). Головка и тело хвостатого ядра явля5 ются передне5боковой стенкой передне5 го рога и средней части бокового же5 лудочка (рис. 6.5, 6.6, 6.7). Хвост явля5 ется крышей нижнего рога бокового же5 лудочка. На фронтальных срезах (рис.
он виден в виде маленького
250 6  ядра и экстрапирамидная система
ядра и экстрапирамидная система
Corpus amygdaloideum
 6.1 Топографические взаимоотно* шения
6.1 Топографические взаимоотно* шения  ганглиев.
ганглиев.
 nuclei
nuclei 
\
 6.2 Отношениия базаль* ных ганглиев к желудочковой системе (вид сбоку).
6.2 Отношениия базаль* ных ганглиев к желудочковой системе (вид сбоку).
округлого образования серого вещества над нижним рогом бокового желудоч5 ка (рис. 6.7). Рострально хвост почти достигает миндалевидного ядра (см. рис. 6.2).
Скорлупа подобно раковине покры5 вает передние отделы бледного шара, будучи отделенной от него слоем мие5
линизированных волокон — боковой медуллярной пластинкой бледного ша5 ра. Латерально, скорлупа отграничена от ограды наружной капсулой (capsula externa). В свою очередь ограда отгра5 ничена от глубинных отделов коры ост5 ровка крайней капсулой (capsula extrema) (рис.  Некоторые из ее волокон
Некоторые из ее волокон
являются ассоциативными, связывая слуховую область височной доли с дви5 гательной и премоторной корой.
О функциях ограды почти ничего неизвестно. Предполагается, что, подоб5 но миндалевидному ядру, она не явля5 ется частью экстрапирамидной систе5 мы. Поэтому в дальнейшем об этих структурах в данной главе упоминаться не будет.
Экстрапирамидная система
Экстрапирамидная система состоит из следующих структур серого вещества: хвостатого ядра, скорлупы, бледного шара, субталамического ядра, черной субстанции, и красного ядра. Термин
экстрапирамидная система кратко оп5 ределен в главе о двигательной систе5 ме — там, где говорится об экстрапи5 рамидных путях, регулирующих спи5 нальную двигательную активность (гла5 ва 2).
Краткий экскурс в филогенез экст5 рапирамидной системы поможет нам в понимании ее функций.
Низшими центрами системы явля5 ются спинной мозг и примитивный ап5 парат ретикулярной формации в по5 крышке среднего мозга. В процессе эво5 люции у животных палеостриатум (бледный шар) начинает эти центры регулировать. В дальнейшем у высших млекопитающих и особенно у человека, развивается неостриатум. Его размеры соотносятся со степенью развития моз5 говой коры. Как правило, низшие цен5 тры подчиняются высшим. Это означа5 ет, что у низших животных низшие центры без особого напряжения зани5 маются регуляцией мышечного тонуса и в значительной мере автоматической регуляцией движений. Но чем выше по филогенетической шкале стоит живот5 ное, тем менее оно способно к компен5 сации нарушений или выпадений вы5 сших центров.
Экстрапирамидная система |
251 |
По мере формирования мозговой коры, филогенетически более старые двигательные центры (палеостриатум и неостриатум) все более подпадают под контроль новой двигательной систе5 мы — пирамидной системы. Тем не ме5 нее, большинство млекопитающих, на5 пример кошки, все еще способны в не5 которой степени передвигаться и даже бегать после удаления моторной коры. Однако для человека интактность пи5 рамидной системы имеет решающее значение, хотя даже больной со спасти5 ческим параличом конечности может выполнять некоторые непроизвольные, т. н. ассоцииированные движения. Наши знания о связях базальных ганглиев друг с другом и другими ядерными образованиями экстрапирамидной сис5 темы все еще недостаточны. Поэтому далее будут обсуждаться только основ5 ные афферентные и эфферентные про5 водящие пути.Полосатое тело является центром высшего порядка среди струк5 тур, составляющих экстрапирамидную двигательную систему. Оно получает импульсы из многочисленных отделов мозговой коры, особенно из двигатель5 ных зон лобной области включая поля 4, 6а и 6а. Эти афферентные волокна расположены в соматотопическом по5 рядке, идут ипсилатерально и, вероятно, являются тормозными. По5видимому, стриокортикальные связи не существу5 ют. Другая система афферентных воло5 кон с прямой передачей импульсов дос5 тигает полосатое тело из таламического центромедианного ядра. Очевидно, эти волокна являются активирующими. Как из хвостатого ядра, так и из скорлупы основные афферентные волокна идут к латеральным и медиальным сегментам бледного шара, разделенным внутрен5 ней медуллярной пластинкой. Вероятно, прямых связей между мозговой корой и бледным шаром не существует. Од5 нако имеется мнение, что существуют ипсилатеральные связи из коры к чер5
252 б Базальные ядра и экстрапирамидная система
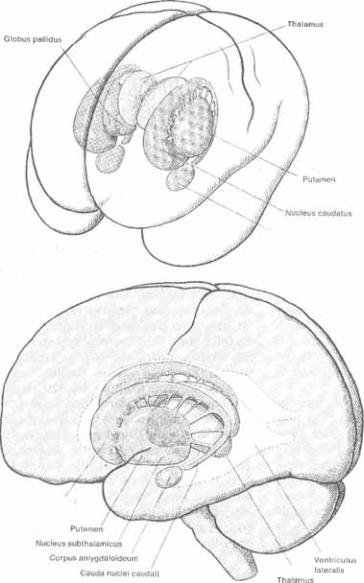
 63 Базальные
63 Базальные  сбо* ку). X и XX обозначают уровень гори* зонтальных срезов через правое (X) и левое (XX) полушария, изображен* ных на Рис. 6.4;
сбо* ку). X и XX обозначают уровень гори* зонтальных срезов через правое (X) и левое (XX) полушария, изображен* ных на Рис. 6.4;  вертикальные линии
вертикальные линии  обознача* ют срезы во фронтальной плоскости, изображенные на Рис.
обознача* ют срезы во фронтальной плоскости, изображенные на Рис. 
 nuclei
nuclei 
 corporis callosi
corporis callosi
Corpus
 nuclei caudati Plexus
nuclei caudati Plexus 
 lateralis
lateralis
superioret inferior
Corpus
\
 6.4 Горизонтальный срез через базальные ганглии.
6.4 Горизонтальный срез через базальные ганглии.
ной субстанции, красному ядру, субта5 ламическому ядру и ретикулярной фор5 мации (рис. 6.9).
Помимо получения афферентных корковых волокон, о чем пойдет речь далее, хвостатое ядро и скорлупа имеют двусторонние связи с черной субстан5 цией.
Афферентные нигростриальные во5 локна считаются  уменьшающими тормозную функцию
уменьшающими тормозную функцию
полосатого тела. С другой стороны, стрионигральный тракт является ГАМК$
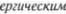 (ГАМК
(ГАМК 
 ляная кислота) и обладает тормозным эффектом на дофаминергические ни5 гро5стриальные нейроны. Это замкну5 тая система с обратной связью (см. рис. 6.9). ГАМК5ергические нейроны стрио5 нигральных волокон, вероятно, подав5 ляют нисходящие, скорее всего дофа5 минергические, нейроны черной суб5 станции, которые регулируют мышеч5 ный тонус через гамма5нейроны (Has5
ляная кислота) и обладает тормозным эффектом на дофаминергические ни5 гро5стриальные нейроны. Это замкну5 тая система с обратной связью (см. рис. 6.9). ГАМК5ергические нейроны стрио5 нигральных волокон, вероятно, подав5 ляют нисходящие, скорее всего дофа5 минергические, нейроны черной суб5 станции, которые регулируют мышеч5 ный тонус через гамма5нейроны (Has5
Все прочие эфферентные волокна
Экстрапирамидная система |
253 |
Corpus 
 lateralis
lateralis
Caput nuclei  Capsula interna
Capsula interna
Insula
Septum 
N.  olfactorius)
olfactorius)
 6.5 1*й фронтальный срез.
6.5 1*й фронтальный срез.
Corpus callosum
Septum pellucidum
Corpus nuclei caudati
Ventriculus lateralis
Capsula interna
Commissura anterior
Insula
Claustrum
Putamen
Area olfactoria
Chiasma opticum
Recessus opticus ventriculi tertii
 6.6 Горизонтальный срез через ба* зальные ганглии.
6.6 Горизонтальный срез через ба* зальные ганглии.
полосатого тела проходят через меди5 альные отделы бледного шара. Они об5 разуют довольно значительные пучки волокон, имеющих окончания на мно5 гих ядрах. Один их этих пучков назы5 вается чечевичная петля 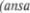
 Ее волокна начинаются в передних отделах медиального сектора бледного шара и идут вентромедиально вокруг задней ножки внутренней капсулы к та5 ламусу и гипоталамусу и, реципрокно,
Ее волокна начинаются в передних отделах медиального сектора бледного шара и идут вентромедиально вокруг задней ножки внутренней капсулы к та5 ламусу и гипоталамусу и, реципрокно,
к субталамическому ядру. После пере5 креста они контактируют с ретикуляр5 ной формацией среднего мозга, откуда начинается цепь нейронов, образующих ретикулоспинальный тракт (нисходя5 щая ретикулярная формация), заканчи5 вающийся на нейронах передних рогов.
Основной массив эфферентных во5 локон бледного шара идет в зрительный бугор как часть нескольких регулирую5 щих систем обратной связи. Этот пал5
254 6 Базальные ядра и экстрапирамидная система
 6.7 3*й фронтальный срез.
6.7 3*й фронтальный срез.
Рис. 6.8 4*й фронтальный срез.
Corpus 
Fornlx
Corpus nuclei caudati
Plexus 
 lateralis
lateralis
Capsule interna
Globus  Nucleus
Nucleus  Cauda nuclei caudati
Cauda nuclei caudati
Hippocampus, Cornu inferius ventriculi lateralis
Tractus (Fasciculus) opticus Fasciculus 
Nucleus corporis mamillaris
 corporis callosum
corporis callosum
Cornu posterius ventriculi lateralis V. cerebri 
Nucleus colliculi inferioris Substantia grisea centralis Aquaeductus cerebri

 superior
superior
—Cerebellum Lemniscus 
Pedunculus cerebellaris 
лидоталамический пучок также называ5
 fasciculus thalamicus или поле Фореля
fasciculus thalamicus или поле Фореля
 Большинство его волокон заканчи5 вается в вентроантериальном (VA) яд5 ре и вентрооральном переднем ядре (V. о. а) зрительного бугра (см. рис. 55). Ядро VA проецируется на корковое поле
Большинство его волокон заканчи5 вается в вентроантериальном (VA) яд5 ре и вентрооральном переднем ядре (V. о. а) зрительного бугра (см. рис. 55). Ядро VA проецируется на корковое поле 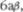 а ядро V. о. а — на корковое
а ядро V. о. а — на корковое
поде  (см. рис. 5.S). Волокна из зуб5 чатого ядра мозжечка заканчиваются в заднем вентрооральном ядре (V. о. р. — см. рис. 55) зрительного бугра, проеци5 рующемся в 4 поле коры. Все эти та5 ламокортикальные пути передают им5 пульсы в обоих направлениях. В коре таламокортикальные пути синапсируют
(см. рис. 5.S). Волокна из зуб5 чатого ядра мозжечка заканчиваются в заднем вентрооральном ядре (V. о. р. — см. рис. 55) зрительного бугра, проеци5 рующемся в 4 поле коры. Все эти та5 ламокортикальные пути передают им5 пульсы в обоих направлениях. В коре таламокортикальные пути синапсируют
с кортикостриальными нейронами и об5 разуют раз ичаые реверберирующие кру5 ги.
Существование различных рецип5 рокных таламокортикальных связей предполагает само собой разумеющим5 ся существование реверберирующих кругов, облегчающих или подавляющих
Экстрапирамидная система |
255 |
активность определенных корковых двигательных полей.
Дополнительными кругами являют5
ся:
1. 
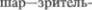 ный бугор (ядро V. о. а.
ный бугор (ядро V. о. а.
2. Хвостатое 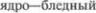
Fasciculus
Ansa 
nigra Nucleus 
 зубчатого
зубчатого 
Tractus rubrospinaiis
Tractus reticulospinalis
 6.9 Связи экстрапирамид* ной системы
6.9 Связи экстрапирамид* ной системы  Hassler).
Hassler).
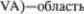
256 б Базальные ядра и экстрапирамидная система
тельный бугор (ядро
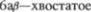 ядро.
ядро.
3. 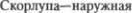 часть бледного
часть бледного
 активирующая
активирующая  ядро зри5 тельного бугра (получающее допол5
ядро зри5 тельного бугра (получающее допол5
нительные импульсы 
 формного
формного 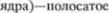 тело.
тело.
4.Наружная часть бледного  тивирующая ретикулярная система— таламические интраламинарные яд5
тивирующая ретикулярная система— таламические интраламинарные яд5
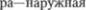 часть бледного шара (не доказано).
часть бледного шара (не доказано).
5.Зубчатое ядро 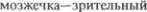 бугор (ядро V. о. р.
бугор (ядро V. о. р. 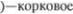 поле
поле 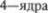 моста (или: красное ядро5
моста (или: красное ядро5
центральный путь 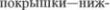 няя
няя  ядро мозжечка (см. рис. 4.5 и 4.6).
ядро мозжечка (см. рис. 4.5 и 4.6).
Нисходящие волокна базальных ганг5 лиев относительно малочисленны по сравнению с паллидоталамическими путями и достигают спиннного мозга с помощью нейрональных цепей. Это заставляет предположить, что основной задачей базальных ганглиев является контроль и регуляция активности дви5 гательных и премоторных корковых по5 лей путем различных реверберирующих кругов, в результате чего произвольные движения становятся плавными и не5 прерывными. Реверберирующие круги полосатого тела дополняются мозжеч5 ковой, вестибулярной и проприоцептив5 ной системами.
Как уже упомянуто в главе 2, пи5 рамидный тракт начинается в сенсомо5 торной коре (поля 4, 6, 3, 1 и 2). В этих же корковых полях берут начало экстрапирамидные двигательные пути: кортикостриарные, кортикорубральные, кортиконигральные, кортикоретикуляр5 ные и др., идущие к двигательным яд5 рам черепных нервов и спинальным двигательным нейронам через нисходя5 щие цепи нейронов. Большинство этих проекций двигательной коры проходит через внутреннюю капсулу. Следова5
тельно, поражения внутренней капсулы обычно прерывают не только волокна пирамидного тракта, но и экстрапира5 мидные пути. Этот перерыв является причиной  Небольшая часть этих экстрапирамидных волокон, веро5 ятно, спускается по наружной капсуле, что может объяснить, почему при спас5 тической гемиплегии вследствие гема5 томы внутренней капсулы парализован5 ная конечность способна выполнять не5 которые произвольные движения.
Небольшая часть этих экстрапирамидных волокон, веро5 ятно, спускается по наружной капсуле, что может объяснить, почему при спас5 тической гемиплегии вследствие гема5 томы внутренней капсулы парализован5 ная конечность способна выполнять не5 которые произвольные движения.
Специфические функции отдельных ядер экстрапирамидной системы до конца не ясны. Полученные в результате стереотаксических вмешательств новые данные привели лишь к условному объ5 яснению. Выявление конкретных выпа5 дений в результате повреждения одного или другого образования не позволяет с уверенностью сделать вывод, что дан5 ное подкорковое ядро является единст5 венным центром, отвечающим за поте5 рянную функцию. Представляется более вероятным, что поражение подкорково5 го ядра или его связей вызывает дис5 координацию обычно гармоничного взаимодействия различных компонен5 тов системы и что клиническая симп5 томатика определяется характером этой дискоординации.
Симптомы поражения  пирамидных образований
пирамидных образований
Основными симпомами экстрапира5 мидных поражений являются наруше5 ния мышечного тонуса (дистония) и расстройства непроизвольных движе5 ний  гипокинезия, акине$
гипокинезия, акине$
 которые во время сна отсутствуют. Можно выделить два клинических синдрома. Первый характеризуется со5 четанием гиперкинезии и гипотонии и
которые во время сна отсутствуют. Можно выделить два клинических синдрома. Первый характеризуется со5 четанием гиперкинезии и гипотонии и
вызывается поражением неостриатума. Второй проявляется сочетанием гипоки$ незии и гипертонии, или ригидности, и вызывается поражением черной суб5 станции. С него мы и начнем.

Синдром 
Этот синдром — классическое проявле5 ние дрожательного паралича или бо$ лезни Паркинсона. В результате дегене5 ративных изменений происходит поте5 ря меланин5содержащих нейронов чер5 ной субстанции и дофаминергических нейронов полосатого тела. Обычно про5 цесс является двусторонним. При од5 носторонней потере нейронов клиниче5 ские признаки возникают на противо5 положной стороне тела.
В случае дрожательного паралича де5 генерация наследственно обусловлена. Однако схожая утрата нейронов черной субстанции может быть обусловлена иными причинами. В этом случае воз5 никший дрожательный паралич назы5 вают 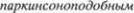 синдромом
синдромом
или паркинсонизмом. Если он является отдаленным последствием летаргиче5 ского энцефалита, его определяеют как
паркинсонизм.
Синдром включает в себя наряду с дру5 гими признаками вегетативные нару5 шения (гиперсаливацию, себоррею ли5 ца), а также глазодвигательные кризы и расстройства аккомодации. Среди других возможных причин паркинсо5 низма следует назвать церебральный атеросклероз, тиф, сифилис мозга, пер5 вичное или вторичное поражение сред5 него мозга опухолью или травмой, ин5 токсикации окисью углерода, марган5 цем, фосгеном или другими вещества5 ми, а также длительный прием лекарств фенотиазинового ряда или резерпина.
Дрожательный паралич
Известен также под именем болезни Паркинсона и характеризуется тремя основными симптомами: акинезией, ри$ гидностью и тремором.
 При акинезии наблюда5 ется медленное снижение подвижности больного. Постепенно угасают все ми5
При акинезии наблюда5 ется медленное снижение подвижности больного. Постепенно угасают все ми5
Экстрапирамидная система |
257 |
мические и экспрессивные движения и все ассоциированные движения. Начало движения, например ходьбы, становит5 ся крайне затрудненным. Вначале боль5 ной должен проделать несколько корот5 ких шажков на цыпочках. Начав дви5 жение, он не может внезапно остано5 виться. Прежде чем сделать остановку, больной вынужден сделать несколько дополнительных шагов вследствие за5 медленной контриннервации. Эта про5 долженная активность называется 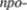
пульсией, ретропульсией или
пульсией, в зависимости от направления движения последних шагов. Лицо ста5 новится маскообразным, поскольку иг5 ра мимической мускулатуры застывает 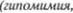
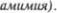 Лишь глаза со5 храняют свою
Лишь глаза со5 храняют свою  Вместо движения головы больной может лишь глазами смотреть в указанном направ5 лении. Речь становится монотонной и дизартричной, частично вследствии ри5 гидности и тремора языка. Наконец, все тело находится в одеревеневшем состоя5 нии антефлексии; все движения крайне медленны и незавершены. Больной из5 бегает любых ненужных движений. Он не размахивает руками при ходьбе. Все сопутствующие мимические и прочие движения носящие индивидуальный ха5 рактер, отсутствуют.
Вместо движения головы больной может лишь глазами смотреть в указанном направ5 лении. Речь становится монотонной и дизартричной, частично вследствии ри5 гидности и тремора языка. Наконец, все тело находится в одеревеневшем состоя5 нии антефлексии; все движения крайне медленны и незавершены. Больной из5 бегает любых ненужных движений. Он не размахивает руками при ходьбе. Все сопутствующие мимические и прочие движения носящие индивидуальный ха5 рактер, отсутствуют.
Ригидность: В противоположность спастическому  мышечного тонуса, ригидность может ощущаться в разгибателях как вязкое воскоподобное сопротивление всем пассивным движе5 ниям. Мышцы не расслабляются. При пассивных движениях можно ощутить ступенеобразное снижение тонуса в мышцах антагонистах (феномен зубча$ того
мышечного тонуса, ригидность может ощущаться в разгибателях как вязкое воскоподобное сопротивление всем пассивным движе5 ниям. Мышцы не расслабляются. При пассивных движениях можно ощутить ступенеобразное снижение тонуса в мышцах антагонистах (феномен зубча$ того  Если у лежащего больного приподнять голову и резко ее отпустить, она не падает сразу, а медленно опус5 кается на подушку (тест падающей го$ ловы). Проприоцептивные рефлексы в отличие от спастического состояния не повышены и никаких патологических
Если у лежащего больного приподнять голову и резко ее отпустить, она не падает сразу, а медленно опус5 кается на подушку (тест падающей го$ ловы). Проприоцептивные рефлексы в отличие от спастического состояния не повышены и никаких патологических