

Реализация мониторинга:
● Роспотребнадзор – федеральная служба по надзору в сфере защиты прав потребителей и благополучия человека.
●Министерство охраны природы и т.д.
Цель мониторинга: информац-ое обеспечение по состоянию прир.
1.Глобальный( межгосударст-ый в пределах всех Земли) Элементы: системы биосферных заповедников( стабильное финансирование, отсчет в глобальн-ой сист.мониторинга)
2.Национальный( в пределах гос-ав) 3.Региональный 4.Локальный уровень
● Документы, к-ые определяют экологическую стандартизацию:
Стандарт – нормативно-технический документ, к-ый устанавливает комплекс норм, требований, обязательных для исполнения.
ГОСТ в 1977г. 140001-76 : система стандартов в обл. охраны природы и улучшении
8 томов: 0 – основные понятия, положения 1-7: гидро-, атмо-, литосферы, флоры, фауна, недры
ООПТ(особо охраняемые прир. тер-рии): участки земли, водной поверх-ти и воздушного пространства над ними, где расположены природ. комплексы, имеющие особое природоохранное, научное, культурное, эстетическое, рекреационное и оздоровительное значение и изъятые решением органов гос-ой власти полностью или частично из хоз-ого использования, и для к-ых установлен тот или иной режим охраны.
Законодательная база
1.Закон об ООПТ – федеральный закон № 33-93 1995
2. 2002 г. закон об охране окр.среды № 7-93
1)Заповедник.
- Биосферные (международные): резерваты ЮНЕСКО
- Государственные: абсолютные (Югыд-Ва), сукцессионные (Басеги), резерватные (Ильменский)
Цель: охрана естественных природ.комплексов и , как следствие этой цели, запрет на любую хоз.деятельность. для создания буфера любой заповедник имеет границы.
На тер-рии возможно проведение мониторинга. – региональные -локальные
2)Государственный природный заказник. Нет задачи полной охраны. Запрещается определенные виды хоз.деятельность. Должны быть прописаны правила!!!Ладшафтный; минералогический; биологический; палеонтологический; гидрологический; геологический.
3)Национальные парки(открытого, курортного и полузакрытого типов, и с заповедным режимом); 2 части: - особо охраняемая территория + хоз.часть( где зарабатываются деньги, чтобы охранять эту тер-рию)
4) Природные парки: организация рекреационных тер-рий, на к-ых имеются охраняемые участки
5)Памятник природы- любой объект, к-ый уникальный в науч, историческом отношении( ландшафтные, гидрологические, геологические, ботанические, археологические)
6)Ботанические сады и дендросады Зоопарки , аквариумы , банк семян: сохранение и база для восстановления природной П-ции и возврат ее в естественную среду – имбридинг (зубр: вливали свежую кровь, т.е. спаривали с бизонами).
Заповедники России: Баргузинский заповедник(1917), Астраханский, Кавказский, Большой Арктический, Командорский, Остров Врангеля.
Пермский край: Басеги (шиверекия подольская, европейский хариус, беркут, сапсан, орлан-белохвост,филин, черный аист ) Вишерский( беркут, орлан-белохвост, скопа) . Заказник: Предуралье. Ботанический сад имени Генкеля, Липовая гора, Сосновый бор…
Красная книга(1963) – юридический документ, к-ый устанавливает статус редких и находящихся под угрозой исчезновения видов раст.и жив; аннотированный перечень видов с указанием прошлого и современного распространения, численности и причин ее сокращения, особенностей воспроизводства, уже принятых и необходимых мер охраны живых организмов. Переиздается каждые 5л.
Категории: 1.Исчезнувшие ( несуществующие) 2.Исчезнувший в дикой природе( сущ-ют за счет разведения в неволе) 3.Находится в критическом состоянии( высокий риск вымирания в дикой прир.в недалеком будущем
- вероятность вымирания сост-ет 50% и более в теч.10 лет или 3х поколений
-насчитывается менее 250 особей этого вида
- менее 50ти особей способны дать потомство
- за последние 10лет числ-ть уменьшилась белее чем на 80%
4.Находится под угрозой исчезновения (имеют высокий риск вымирания в дик.прир.в недалеком буд-ем) вероятность – 20% в теч.20лет или 5ти поколений 5.Уязвимые( имеют риск вымирания в перспективе: риск 10% в теч.100 лет) 6.Требуют сохранения (прямой угрозы вымир-я нет, но нужна специальная программа по сохран-ю) 7.Сущест-ет возможная угроза исчезновения( в наст.вр. видам ничего не угрожает) 8.Не нуждаются в охране 9.Данные отсутствуют 10.Без оценки ( с т.з. вымирания вид не оуценивается)
Международная красная книга ● 1996- Красная книга Урала● 2008- Красная книга Пермского края
Категории в книге: 0: вероятно исчезнувшие 1:в критическом состоянии 2: в опасном состоянии
3:уязвимые ( редкие)
Статусы КК: – Красные страницы – исчезающие виды – Белые – редкие – Желтые – под угрозой исчезновения – Серые - неопределенные – Зеленые – численность восстановлена
Международные соглашения по охране видов
Ответ-ть за сохранение и воспроизводство объектов не несут физические и юридические лица, осущ-щие деят-ть на тер-рии, на к-ой обитают эти виды.
1)Международная конвенция о торговле видами дикой фауны и флоры, наход-ся под угрозой исчез-ия 2)Конвенция по охране мигрирующих видов 3)Конвенция по регулированию добычи диких видов жив-ых 4)Конвенция по рыбной ловле 5)Международная конвенция по защите птиц
Самая эфф-ная мера по сохранению редких и исчезающих видов – сохранение их местообитания, к-ое м.б. достигнуто, в частности, организацией ООПТ.
Вымершие птицы: дронт 1680-1750; Эпиорнис – страусоподобная птица 450 кг; бескрылая гагарка( пингвин Сев.полушария)
Млекопитающие: Европейский тур; квага( полулошадь полузебра); Тарпан; Стеллерова корова; сумчатый волк.
За последние 30-40лет: золотая жаба, занзибарский леопард, пиренейский горный козел, западноафриканский черный носорог.
Определение группы сцепления и локализация генов в группе сцепления. Локализация летальных генов.
Закон сцепления гласит: сцепленные гены, расположеные в одной хромосоме, наследуются совместно (сцепленно).
Описанный Менделем механизм наследования работает только при полном доминировании и только в том случае, если эти признаки контролируются генами, лежащими в разных хромосомах, то есть, не передаются сцеплено. Совместное наследование генов, ограничивающее их свободное комбинирование, называется сцепленным наследованием. Группу сцепления образуют все гены, локализованные в одной хромосоме, эта группа наследуется независимо от других групп сцепления. Число групп сцепления равно гаплоидному набору хромосом.
Число групп сцепления генов у каждого вида животных и растений должно соответствовать гаплоидному числу хромосом данного вида;
Гены в каждой группе сцепления расположены в определенной последовательности; иначе говоря, каждая хромосома в отношении расположения генов имеет свою топографию.
Совпадение числа групп сцепления с гаплоидным числом хромосом служит одним из доказательств того, что именно хромосомы являются материальными носителями наследственной информации.
Принцип определения принадлежности гена к той или иной группе сцепления сводится к тому, чтобы установить характер наследования определяемого гена по отношению к одному или двум другим генам, находящимся уже в известной группе сцепления.
Предположим, что нам известно по одному рецессивному гену в каждой из 10 групп сцепления у кукурузы (b, с, d, е, f и т. д.) и необходимо определить, к какой из групп сцепления относится рецессивный ген а. Так как этот ген может относиться лишь к какой-то одной группе сцепления, то с генами из остальных 9 он должен показывать независимое наследование. Так, при скрещивании двух генотипов A/A d/d x a/a D/D в F2 будет расщепление 9А—D— : 3А—dd : 3aaD— : 1aadd, а в анализирующем скрещивании 1AaDd : 1Aadtf : 1aaDd : 1aadd. Однако с геном одной из 10 групп сцепления, допустим с геном c, ген а обнаружит сцепление. Если сцепление окажется полным Ac/Ac x aC/aC, то при скрещивании в F2 будет наблюдаться расщепление 2АсаС : 1AcAc : 1aCaC, а в анализирующем скрещивании 1ACac : 1aCac.
В указанных примерах мы разобрали два крайних случая — полное сцепление с геном одной из групп сцепления и независимое наследование с генами остальных групп. Однако случай полного сцепления встречается в таких опытах крайне редко. Как правило, устанавливается неполное сцепление исследуемого гена с геном одной из групп сцепления. В данном случае при анализирующем скрещивании Ac/aC x ac/ac в потомстве обнаруживаются все четыре класса генотипов Аасс, ааСс, АаСс и аасс, но в неравных количествах: сочетаний, характерных для исходных родительских форм (А—сс и ааС—), всегда будет больше, чем новых сочетаний (А—С— и аасс). В F2 в данном случае особей аасс должно быть меньше, чем их ожидалось бы при независимом наследовании (9:3:3:1).
Изложенные выше методы применяются для определения группы сцепления у аутосомных генов. Гены, расположенные в половой хромосоме, как правило, показывают наследование, сцепленное с полом. Поэтому установление группы сцепления для таких генов несколько упрощается.
У объектов, группы сцепления которых еще не все маркированы мутантными генами, эта работа усложняется. В этом случае гены, которые при испытании не попадают ни в одну из известных уже групп сцепления, могут быть отнесены к новой, еще не известной группе сцепления. Обычно специально для работы по определению групп сцепления создают генетические линии, у которых каждая из хромосом маркирована хотя бы одним мутантным геном. Скрещивание с одной такой линией позволяет сразу определить группу сцепления для изучаемого гена.
Однако при определении группы сцепления только генетическими методами не всегда можно сказать, какая конкретная пара гомологичных хромосом аналогична соответствующей группе сцепления. Для этого требуются дополнительные цитогенетические исследования.
Локализация гена После определения группы сцепления, к которой принадлежит ген, переходят к следующему этапу анализа и устанавливают место гена в группе сцепления. Локализация гена осуществляется путем учета результатов кроссинговера. Иногда для локализации гена привлекают и цитологические методы.
Для нахождения местоположения локуса гена в хромосоме необходимо производить скрещивание особей таким путем, чтобы место определяемого гена при кроссинговере было третьей точкой, как это было нами уже изложено при объяснении двойного кроссинговера. Маркировка трех локусов в хромосоме, или, как иногда называют, трех точек, совершенно необходима для определения порядка линейного расположения генов и расстояния между ними.
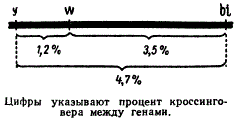
Схема определения локуса в гена в хромосоме
В рассмотренном ранее примере с дрозофилой процент кроссинговера между генами y и w равен 1,2, а между генами w и bi — 3,5. Но по этим показателям мы еще не можем сказать, где находится ген y, слева или справа от гена w; нельзя ничего сказать и о положении гена w по отношению к bi. И только определив процент перекреста между третьей парой генов y и bi в данном случае (4,7%), можно прийти к заключению, что ген w должен находиться между генами y и bi.
Таким образом, с помощью кроссинговера можно определить группу сцепления и место гена в ней. При этом внутри самого гена перекрест обычно не обнаруживается, в силу этого некоторые генетики склонны были определять ген как единицу перекреста, а точнее — как предельную единицу, не разделяемую далее перекрестом.
Поскольку ген занимает определенное место в группе сцепления, то это позволяет генетикам изучать топографию расположения генов в каждой хромосоме и представить это в виде генетических карт хромосом.
Анеуплоиды систематически появляются в скрещиваниях полиплоидов (2n×4n, 3n×3n и т. д.), при межвидовой гибридизации, ни могут возникать в результате нарушений в мейозе, например при нерасхождении хромосом. Это - трисомики (2n+1), моносомики (2n-1), тетрасомики (2n+1+1), нуллисомики (2n-2). У анеуплоидов избыточные или недостающие хромосомы принадлежат к одной гомологической паре. Диплоидные растения (2n) являются нормальными дисомиками, так как все хромосомы у них парные. Анеуплоидные серии используют для определения группы сцепления у растений.
а. Определение группы сцепления с помощью трисомиков
Трисомиков (2n+1) для генетического анализа обычно выделяют в потомстве триплоидных растений, где наблюдается большое разнообразие и наибольшая частота трисомиков по разным хромосомам, или их получают в скрещиваниях между диплоидными и тетраплоидными формами.
Трисомики у растений обычно менее плодовиты и жизнеспособны, чем дисомики. В мейозе у них образуются либо тривалент, либо бивалент и унивалент, либо три унивалента. Это зависит от вида растения и дополнительной хромосомы трисомика. Например, у кукурузы чаще встречаются триваленты, а у сахарной свеклы и томатов - бивалент и унивалент. Остальные хромосомы трисомика конъюгируют нормально, образуя биваленты. При распределении хромосом в мейозе у трисомиков образуются два типа гамет - n и n+1, причем дополнительная хромосома чаще передается через яйцеклетку, так как пыльцевые зерна с лишней хромосомой, как правило, не прорастают, либо их рост замедлен. Поэтому трисомиков используют в скрещиваниях в качестве материнских форм. Они могут иметь четыре разных генотипа по одному гену - AAA, AAa, Ааа, ааа. Расщепление по генам, локализованным в лишней хромосоме, происходит по тем же законам, что и у эуплоидов, т. е. имеет место как случайное хромосомное, так и случайное хроматидное расщепление с учетом двойной редукции (см. гл. V).
Для локализации гена с помощью этого метода последовательно скрещивают диплоидные формы, в генотипе которых содержится локализуемый ген, с набором трисомиков по всем хромосомам. В одном из этих скрещиваний, которое называется "критическим", появляется новый гетерозиготный трисомик. Для локализации рецессивной мутации диплоидную форму (аа) скрещивают с триплоидом, имеющим генотип AAA. Для локализации доминантной мутации используют трисомики с рецессивным генотипом (ааа). В первом поколении в любом из этих скрещиваний возникают новые трисомики с генотипами ААа или Ааа соответственно. Их выделяют либо по фенотипу, если они отличаются от диплоидов, либо цитологически. Затем самоопыляют или проводят анализирующее скрещивание с рецессивным диплоидом. Ниже приведены схемы скрещиваний для получения гетерозиготных трисомиков.

При случайном хромосомном расщеплении в потомстве от самоопыления трисомиков из F1 возникнут следующие расщепления:

При случайном хроматидном расщеплении вероятность выщепления рецессива в F2 от самоопыления трисомика с генотипом ААа значительно меньше, так как он образует 3 типа гамет (С62) с разной вероятностью: 6АА, 1аа и 8Аа, что приведет к расщеплению 44А:1а.

Аналогично можно рассчитать расщепление в Fa при случайном хромосомном и случайном хроматидном расщеплениях этих трисомиков.
Следует помнить, что результаты реальных скрещиваний редко совпадают с рассчитанными теоретически, так как у трисомика обычно образуется больше микро- и мегаспор с гаплоидным набором хромосом, а пыльца с набором n+1 часто стерильна или мало конкурентоспособна, обычно в потомстве трисомика преобладают дисомики и наблюдается дефицит рецессивов в критическом скрещивании.
Таким образом с помощью трисомиков удается локализовать гены в определенной группе сцепления, метод с успехом применяется на ряде диплоидных видов. Наиболее трудоемкая часть работы при его использовании - создание полной серии трисомиков.
б. Определение группы сцепления с помощью моносомиков и нуллисомиков
Моносомики - организмы с недостатком одной из хромосом (2n-1).
Поскольку у диплоидов моносомики, как правило, нежизнеспособны или мало жизнеспособны, моносомные формы используют для локализации генов лишь у аллополиплоидов, у которых гибридологический метод анализа очень сложен и с его помощью практически невозможно решить эту задачу.
На основе межвидовой гибридизации созданы серии моносомиков по всем хромосомам у аллотетраплоидного табака (Nicotiana tabacum) и у гексаплоидной мягкой пшеницы (Triticum aestivum), которая представляет собой сложный полиплоид (2n = 42). Первые моносомики мягкой пшеницы были получены Сирсом в 1939 г. от гаплоида, спонтанно возникшего среди гибридов F1 при скрещивании сорта пшеницы Чайниз Спринг (2n = 42) с рожью Secale cereale (2n = 14). У этого гаплоида было 28 хромосом (21 +7), и он имел стерильную пыльцу. После опыления его пыльцой пшеницы сорта Чайниз Спринг было получено тринадцать жизнеспособных семян. Среди растений, выросших из этих семян, пять оказались моносомиками, образующими в мейозе 20 бивалентов и один унивалент (2011+11). Среди остальных - шесть сказались дисомиками, нуллисомиками и моносомиками с различными аномалиями в мейозе. В дальнейшем Сирсом была создана полная серия моносомиков пшеницы сорта Чайниз Спринг, которая используется для получения новых серий моносомиков на других сортах. Для этого материнские моносомные растения сорта Чайниз Спринг скрещивают с отцовскими дисомными растениями из данного сорта. В потомстве от этих скрещиваний путем цитологического анализа отбирают моносомные растения, получившие свою единственную хромосому от отца. Эти растения вновь скрещивают с отцовским сортом и такой бэккросс повторяют не менее шести раз, поэтому новые моносомики становятся идентичными отцовскому сорту.
Для определения
группы сцепления последовательно
скрещивают диплоидные растения,
гомозиготные по локализуемому гену, со
всеми моносомными линиями сорта. В
потомстве от этих скрещиваний цитологически
идентифицируют вновь возникших
моносомиков. Если рецессивная мутация
полностью проявляется в гемизиготном
состоянии, то все моносомики F1 в
скрещивании ![]()
![]() будут
иметь мутантный фенотип и группа
сцепления определяется уже в первом
поколении. Иногда рецессивный
фенотип не проявляется в гемизиготе и
моносомные потомки такого скрещивания
(а) имеют нормальный фенотип. Моносомики,
возникающие в F1 при
скрещивании
будут
иметь мутантный фенотип и группа
сцепления определяется уже в первом
поколении. Иногда рецессивный
фенотип не проявляется в гемизиготе и
моносомные потомки такого скрещивания
(а) имеют нормальный фенотип. Моносомики,
возникающие в F1 при
скрещивании ![]() неотличимы
от дисомиков и также имеют нормальный
фенотип. В этих случаях локализацию
генов проводят на основе расщепления
в F2, получаемого от
самоопыления моносомиков F1,
выявляемых цитологически. В
потомстве моносомика _а рецессивный
фенотип может проявиться либо у
гомозиготных дисомиков
неотличимы
от дисомиков и также имеют нормальный
фенотип. В этих случаях локализацию
генов проводят на основе расщепления
в F2, получаемого от
самоопыления моносомиков F1,
выявляемых цитологически. В
потомстве моносомика _а рецессивный
фенотип может проявиться либо у
гомозиготных дисомиков ![]() ,
либо у нуллисомиков, лишенных этой
хромосомы. В потомстве моносомика А
рецессивный фенотип проявляется только
у нуллисомиков. Моносомики, дисомики и
нуллисомики выщепляются в потомстве
самоопыленных моносомных растений,
образующих два сорта гамет - n и n-1.
Соотношение этих типов гамет и
соответственно, образование моно-, ди-
и нуллисомных растений зависит, главным
образом, от сорта. Так, через материнские
моносомные растения сорта Чайниз Спринг
передается от 70 до 81% гамет с недостающей
хромосомой (в среднем - 75%), а через
отцовские - лишь 4%, благодаря чему в
F2 возникает следующее соотношение
разных типов растений.
,
либо у нуллисомиков, лишенных этой
хромосомы. В потомстве моносомика А
рецессивный фенотип проявляется только
у нуллисомиков. Моносомики, дисомики и
нуллисомики выщепляются в потомстве
самоопыленных моносомных растений,
образующих два сорта гамет - n и n-1.
Соотношение этих типов гамет и
соответственно, образование моно-, ди-
и нуллисомных растений зависит, главным
образом, от сорта. Так, через материнские
моносомные растения сорта Чайниз Спринг
передается от 70 до 81% гамет с недостающей
хромосомой (в среднем - 75%), а через
отцовские - лишь 4%, благодаря чему в
F2 возникает следующее соотношение
разных типов растений.

Если моносомик имеет генотип а, то в F2 возникнет соотношение: 73% растений с нормальным фенотипом (моносомики) и 27% с рецессивным фенотипом (дисомики и нуллисомики). Моносомик с генотипом А в F2 будет давать иное соотношение: 97% нормальных растений (дисомики и моносомики А) и 3% -рецессивных (нуллисомики).
Эти частоты приняты в качестве стандарта при оценке расщеплений в потомстве самоопыленных моносомиков.