11. Внутрішнє обертання та поворотна ізомерія в макромолекулах.
Процес формування конформації.

Класична органічна хімія вважала вільним обертання атомних груп навколо одиночних зв'язків. Будь-які конформації, наприклад, етану Н3С-СН ,, що виникають в результаті внутрішніх поворотів, мають однакову енергію;

зміна проекції на площину, перпендикулярну С - С-зв'язку
кута повороту не вимагає витрати енергії. Деякі з конформаций етану зображені на рис. 3.1.
Однак дослідження термодинамічних властивостей етану та інших з'єднань з одиничними зв'язками, а також структурні дослідження, проведені методами спектроскопії, ЯМР і т. Д., Довели, що внутрішнє обертання майже завжди невільно.

Молекула етану має мінімум енергії в
транс-, або схрещеною, конформації (рис.
3.1, а) і максимум в цис-, або затененной,
конформації. Для повороту навколо
С-С-зв'язку на 120 °, т. Е. Для переходу з
однієї тракс-конформації в іншу, їй
тотожну, потрібно подолати енергетичний
бар'єр, равний 12 200 Дж / моль. Для етану,
а також для інших молекул з осьової
симетрією С3 залежність потенційної
енергії молекули від кута повороту ф
можна наближено представити формулою![]()
Детальна конфірмаційна енергія полімерного ланцюга.

12. Гнучкі та жорсткі полімерні ланцюги. Високоеластичність.
Гнучкість – можливість ланцюгу згортатися довжинами хім.. звязів, валентними кутами між ними та кутами внутр.. обертання
Модель очленованого ланцюгу.

який з’єднує початок та
кінець молекули. За доп. методів стат.
фізики можна знайти, що сер. значення=0,
беруть секредньоквадратичне![]() .
Видно, що макромолекула має обємну
форму. Всі макромолекули мають певну
гнучкість, яка в данній моделі має
шарнірними зєднаннями елементів. міра
гнучкості <R2>.
.
Видно, що макромолекула має обємну
форму. Всі макромолекули мають певну
гнучкість, яка в данній моделі має
шарнірними зєднаннями елементів. міра
гнучкості <R2>.

Модель жорсткого ланцюгу
Персистентна або червоно подібна (для жорстких ланцюгів).

![]() фі – кут між 2 дотичними до 2-х точок
ланцюгу, що розділені між собою контурною
довжиною L. Відомо, що якщо контурна
довжина менше а, то можна вважати ланки
малі і залежні молекулою гнучкості
якщо бильше а, то до моделі твердого
стержня.
фі – кут між 2 дотичними до 2-х точок
ланцюгу, що розділені між собою контурною
довжиною L. Відомо, що якщо контурна
довжина менше а, то можна вважати ланки
малі і залежні молекулою гнучкості
якщо бильше а, то до моделі твердого
стержня.
 За
данною ф-лою оцінюють гнучкість або
жорсткість .
За
данною ф-лою оцінюють гнучкість або
жорсткість .
Високоеластичність – хар. яка, полягає в тому, що при розтягненні молекули розмір в декілька разів за рахунок розгортання ланцюгу. Після зняття напруги знов згортається.

13. Перехід «клубок-глобула».
Хар. макромолекул, яка хар. внутр.. просторову стр. молекули
 Викор.
стат. хар. Вводимо поняття локальної
густини мономерів:
Викор.
стат. хар. Вводимо поняття локальної
густини мономерів:
![]() .
В наслідок обємних взаємодій густина
мономерів може змін. від точки до точки.
Існує просторова кореляція густини.
.
В наслідок обємних взаємодій густина
мономерів може змін. від точки до точки.
Існує просторова кореляція густини.
Клубком назив. такий стан, в якому
флуктуації густини порядку собою
густини.![]() .
.
Радіус

Глобула – такий стан, в якому флуктуації густини багото менше самої густини самої густини і радіус кореляції самої густини багато меньша розміру молекули.
Особливості поверхні глобули.

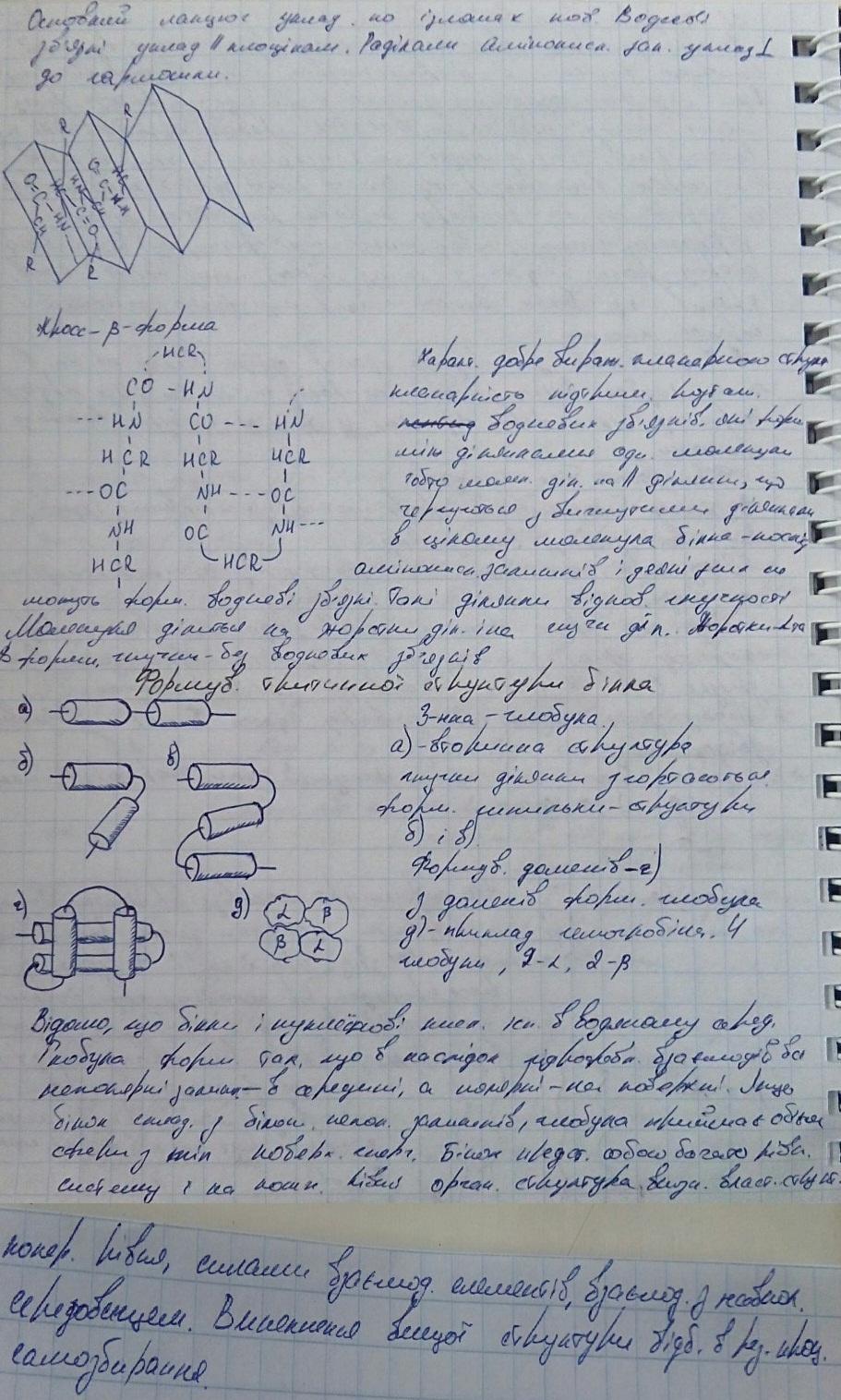
14. Будова білкової глобули.
білкова глобула є ієрархічна структура К-сть конформації для білка обмежена.