

Гліоксилатний цикл
Гліоксилатний цикл локалізований не у мітохондріях, як цикл Кребса, а у спеціалізованих мікротілах-гліоксисомах. На відміну від циклу Кребса, у гліоксилатному циклі бере участь не одна, а дві молекули ацетил-СоА. При цьому ацетил-СоА використовується не для окислення, а для синтезу бурштинової кислоти (рис. 56).

Рис. 56. Гліоксилатний цикл.
З щавлевооцтової кислоти і ацетил-СоА синтезується лимонна кислота, потім утворюються цис-аконітова та ізолимонна, як і у циклі Кребса.
Потім ізолимонна кислота за участі ферменту ізоцитратліази розпадається на гліоксилеву і бурштинову кислоти. Гліоксилева кислота під дією малатсинтетази взаємодіє з другою молекулою ацетил-СоА, що призводить до утворення яблучної кислоти, яка потім перетворюється у фосфоенолпіровиноградну кислоту, а остання – у вуглеводи.
Цей цикл активно функціонує у проростаючому насінні олійних рослин та інших органах, де запасні жири перетворюються у цукри.
Гліоксилатний цикл дає можливість використання запасних жирів, унаслідок розпаду яких утворюються молекули ацетил-СоА. Крім того, на кожні дві молекули ацетил-СоА у гліоксилатному циклі відновлюється одна молекула НАДН2, енергія якої може бути використана на синтез АТР у мітохондріях або в інших процесах.
Пентозофосфатний цикл
У рослинній клітині поряд з розпадом глюкози шляхом гліколізу і у циклі Кребса, який є головним постачальником енергії, відбувається і другий важливий спосіб її перетворення – пентозофосфатний цикл (рис. 57).
У цьому випадку розщепленню глюкози передує утворення її монофосфорного ефіру, а потім нібито відсічення першого вуглецю у ланцюзі. Відповідно до цього пентозофосфатний цикл часто називають апотомічним окисленням на відміну від гліколітичного циклу, який називають дихотомічним за утворення у ньому двох тріоз.
Окислення глюкози пентозофосфатним шляхом відбувається у цитоплазмі клітини, а також у пластидах. Особливо висока активність пентозофосфатного шляху дихання у клітинах і тканинах з активними синтетичними процесами. Розщеплення глюкози, як і при гліколізі, розпочинається з її фосфорилювання. За допомогою ферменту гексокінази та АТР утворюються глюкозо-6-фосфат і АДР. У реакціях пентозофосфатного циклу беруть участь шість молекул глюкози, п’ять із яких регенеруються, а одна окислюється відповідно до рівняння:
![]()
У цьому циклі можна виділити два етапи: а) окислення глюкози; б) регенерація вихідного субстрату.
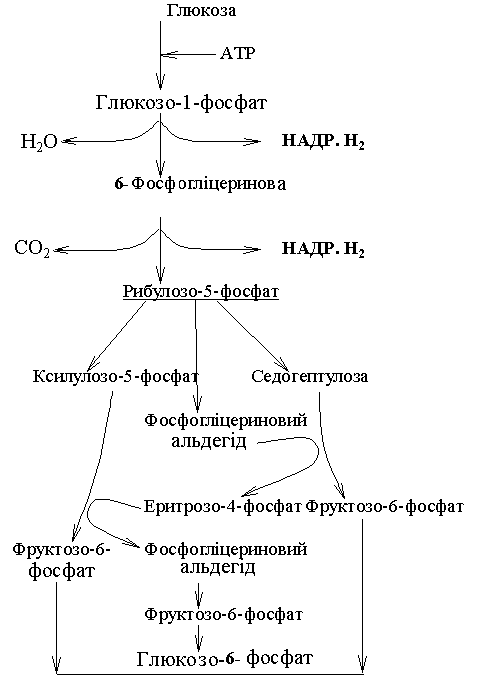
Рис. 57. Пентозофосфатний цикл.
Реакції першого етапу каталізує дегідрогеназно-карбоксилювальна система з трьох ферментів. Спочатку відбувається дегідрування глюкозо-6-фосфату за участю ферменту глюкозо-6-фосфатдегідрогенази, у активній групі якого є НАДФ – акцептор електронів. Утворена 6-фосфоглюконова кислота зазнає окислювального декарбоксилювання і дегідрування до рибулозо-5-фосфату.
Другий етап циклу пов’язаний із регенерацією глюкозо-6-фосфату. Із рибулозо-5-фосфату під дією ізомераз утворюється рибозо-3-фосфат. Потім із двох фосфопентоз шляхом рекомбінації при послідовній участі ферментів транскаталази і трансальдолази утворюється спочатку семивуглецевий цукор седогептулоза і фосфогліцериновий альдегід, потім – еритрозофосфат і фруктозо-6-фосфат. Унаслідок ізомеризації фруктозо-6-фосфат переходить у глюкозо-6-фосфат.
Для кожного оберту підсумкове рівняння пентозофосфатного циклу має вигляд:
![]()
Як випливає з цього рівняння, при повному окисленні однієї молекули глюкозо-6-фосфату утворюється 12 молекул НАДФН2. Окислення 12 пар протонів від НАДФН2 у процесі окислювального фосфорилювання забезпечує синтез 36 молекул АТР, що становить 41,87кДж 36 = 1507кДж/моль і практично не поступається енергетичному виходу гліколітичного шляху дихання.
Окислення глюкози пентозофосфатним шляхом виявлено в різних органах рослини. Воно є головним джерелом утворення пентоз, які використовуються клітиною для синтезу нуклеїнових кислот. Цей цикл також постачає рибозу, яка у формі рибулозодифосфату слугує акцептором вуглекислого газу у темновій фазі фотосинтезу. Сполуки з різною кількістю атомів вуглецю використовуються для багатьох біосинтезів. Так, еритрозо-4-фосфат, крім того, що може сполучатися із ксилулозо-5-фосфатом, утворюючи фруктозо-6-фосфат і фосфогліцериновий альдегід, також може приєднувати фосфоенолпіровиноградну кислоту і давати шикимову кислоту – циклічну сполуку (рис. 58), яка є попередником для синтезу ряду ароматичних сполук (поліфеноли, дубильні речовини, амінокислоти, глікозиди), що відіграють важливу роль у обміні речовин. Одна із циклічних амінокислот – триптофан – є попередником гетероауксину.

Рис. 58. Участь шикимової кислоти у метаболізмі рослинного організму.
Перетворення глюкози пентозофосфатним шляхом регулюється концентрацією НАДР+, яка залежить від синтетичних процесів, де використовується НАДФН2 (синтез амінокислот, білків).
Між гліколізом і пентозофосфатним шляхом перетворення глюкози існує тісний зв’язок, оскільки у них є багато спільних ферментів і проміжних продуктів. Ці два метаболічні шляхи майже в усіх випадках функціонують одночасно і продукти пентозофосфатного перетворення у певних умовах піддаються подальшому використанню в реакціях гліколітичного шляху.
Співвідношення між пентозофосфатним шляхом перетворення глюкози і гліколізом залежить від наявності проміжних сполук (неорганічного фосфату, еритрозо-4-фосфату і 6-фосфоглюконової кислоти).