

М. Р. Тайрова, Н. В. Громова
АНАТОМИЯ И ФИЗИОЛОГИЯ НЕРВНОЙ И СЕНСОРНЫХ СИСТЕМ
(ВОЗРАСТНЫЕ ОСОБЕННОСТИ)
УЧЕБНОЕ ПОСОБИЕ
ОГЛАВЛЕНИЕ
РАЗДЕЛ I. АНАТОМИЯ И ФИЗИОЛОГИЯ НЕРВНОЙ СИСТЕМЫ
|
Глава 1.Общий план строения и физиология нервной системы..........
|
Глава 2. Частная физиология центральной нервной системы.............
|
РАЗДЕЛ II. АНАТОМИЯ И ФИЗИОЛОГИЯ СЕНСОРНЫХ СИСТЕМ (АНАЛИЗАТОРОВ)
|
Глава 1. Общая схема строения анализаторов.........................................
|
Глава 2. Зрительный анализатор................................................................
|
Глава 3. Строение и функции слухового анализатора............................
|
Глава 4. Вестибулярный анализатор..........................................................
|
Глава 5. Обонятельный анализатор...........................................................
|
Глава 6. Вкусовой анализатор.....................................................................
|
Глава 7. Кожный анализатор.......................................................................
|
Литература......................................................................................................
|
Раздел I. Анатомия и физиология нервной системы
Глава 1. Общий план строения и физиология нервной системы
1. 1 Значение нервной системы. Общий план ее строения. Нервная система выполняет в организме человека и высших животных ряд важнейших функций: анализаторную (анализ поступающей по сенсорным системам информации), моторную (организация ответных движений), интегративную (объединение, связь различных сенсорных и моторных структур нервной системы; формирование временных связей, обучение), организацию психических процессов. Кроме того, нервная система регулирует и интегрирует функции внутренних органов (висцеральная функция).
Нервная система в функциональном и структурном отношении делится на периферическую и центральную нервную системы. Центральная нервная система (ЦНС) – совокупность связанных между собой нейронов. Она представлена головным и спинным мозгом. На разрезе головного и спинного мозга различают участки более темного цвета – серое вещество (образовано телами нервных клеток) и участки белого цвета – белое вещество мозга (скопление нервных волокон, покрытых миелиновой оболочкой).
Периферическая часть нервной системы образована нервами (пучками нервных волокон, покрытых сверху общей соединительнотканной оболочкой), нервными узлами, или ганглиями (скопления нервных клеток вне спинного и головного мозга) и рецепторами. Если в составе нерва собраны нервные волокна, передающие возбуждение из ЦНС к иннервируемому органу (эффектору), то такие нервы называют двигательными, центробежными, или эфферентными. Есть нервы, которые образованы чувствительными нервными волокнами, по которым возбуждение распространяется в ЦНС. Такие нервы называют центростремительными, или афферентными. Большинство нервов являются смешанными, в их состав входят как центростремительные, так и центробежные нервные волокна.
Разделение нервной системы на центральную и периферическую во многом условно, так как функционирует нервная система как единое целое.
По функциональным свойствам нервную систему делят на соматическую, или цереброспинальную, и вегетативную. К соматической относят ту часть нервной системы, которая иннервирует опорно-двигательный аппарат и обеспечивает чувствительность нашего тела. К вегетативной – все другие отделы, которые регулируют деятельность внутренних органов (сердце, легкие, органы выделения и др.), гладких мышц сосудов и кожи, различных желез и обмен веществ (обладает трофическим влиянием на все органы, в том числе и на скелетную мускулатуру).
Вегетативная нервная система (ВНС) состоит из двух отделов: симпатического и парасимпатического. Симпатический отдел ВНС обеспечивает адаптацию организма к разнообразным изменениям внешней среды. Причем симпатический отдел в ситуациях, требующих мобилизации всех защитных сил организма (стресс), способствует интенсификации вегетативных процессов: увеличению частоты и силы сердечных сокращений, сужению кровеносных сосудов, замедлению двигательной функции желудочно-кишечного тракта, усилению обмена веществ и т. д. Все симпатические нервы выходят из спинного мозга (от нейронов боковых рогов, расположенных на уровне VIII шейного – II поясничного сегментов) в составе передних корешков и, отделившись от них, идут к пограничному симпатическому стволу. Часть из них прерывается в узлах симпатического ствола, другая часть проходит, не прерываясь. Парасимпатический отдел способствует восстановлению гомеостаза, осуществляет «текущую» регуляцию функций организма. Парасимпатические нервные волокна имеются в составе черепно-мозговых нервов (III пара – средний мозг; VII пара – мост, IX и X пары – продолговатый мозг) и в тазовом нерве – крестцовый отдел спинного мозга. Симпатические волокна имеют значительно более широкое распространение, чем парасимпатические. Симпатические нервы иннервируют фактически все органы и ткани организма; парасимпатические же не иннервируют скелетную мускулатуру, ЦНС, большую часть кровеносных сосудов, матку.
А. Д. Ноздрачев ввел понятие «метасимпатическая» нервная система. Он считает, что это комплекс микроганглионарных образований, расположенных в стенках внутренних органов, обладающих моторной активностью (микроганглии в желудке, кишечнике, мочевом пузыре, сердце, бронхах).
1. 2 Основные этапы развития нервной системы. Последствия неблагоприятных воздействий на организм в критические периоды развития нервной системы. Нервная система начинает формироваться на третьей неделе эмбрионального развития из дорсальной части наружного зародышевого листка (эктодермы). Сначала образуется нервная пластинка, которая постепенно превращается в желобок с приподнятыми краями. Края желобка приближаются друг к другу и образуют замкнутую нервную трубку. Из нижнего (хвостового) отдела нервной трубки образуется спинной мозг, из переднего – все отделы головного мозга: продолговатый мозг, мост и мозжечок, средний мозг, промежуточный мозг и большие полушария.
В головном мозге различают по происхождению, структурным особенностям и функциональному значению три отдела: ствол, подкорковый отдел и кору больших полушарий. Мозговой ствол – это образование, расположенное между спинным мозгом и большими полушариями. К нему относят продолговатый, средний и промежуточный мозг, мост и мозжечок. К подкорковому отделу относят базальные ганглии. Кора больших полушарий является высшим отделом головного мозга.
В процессе развития из переднего отдела нервной трубки образуются три расширения – первичные мозговые пузыри (передний, средний и задний, или ромбовидный). Эту стадию развития головного мозга называют стадией трехпузырного развития.
У трехнедельного эмбриона намечается, а у пятинедельного хорошо выражено разделение поперечной бороздой переднего и ромбовидного пузырей еще на две части, вследствие чего образуется пять мозговых пузырей (стадия пятипузырного развития), которые дают начало всем отделам головного мозга. Мозговые пузыри растут неравномерно. Наиболее интенсивно развивается передний пузырь, который уже на ранней стадии развития разделяется продольной бороздой на правый и левый. На третьем месяце эмбрионального развития сформировано мозолистое тело, которое соединяет правое и левое полушария, а задние отделы переднего пузыря полностью покрывают промежуточный мозг. На пятом месяце внутриутробного развития плода полушария распространяются до среднего мозга, а на шестом – полностью покрывают его. К этому времени все отделы головного мозга хорошо выражены.
Клетки коры и двигательные поводящие пути к моменту рождения ребенка недоразвиты. Количество полушарных извилин, их форма, топографическое положение претерпевают определенные изменения по мере роста организма (наибольшие изменения происходят в течение первых 5 – 6 лет) и лишь к 15 – 16 годам становятся похожими на мозговые структуры взрослого человека. К 18 – 20 годам формирование нервной системы завершается.
Мозг наиболее чувствителен к различным неблагоприятным факторам в критические периоды своего развития, когда наблюдается интенсивная дифференциация нервной системы. Неблагоприятные воздействия на плод в период от 3 до 10 недель развития могут быть причинами формирования грубых пороков развития нервной системы. Если нарушения возникают на более поздних этапах онтогенеза, выраженность дефекта может варьировать в разной степени: от грубого нарушения функции или ее полного отсутствия до легкой задержки темпа развития. Причины возникновения дефектов и нарушений развития разнообразны. Это могут быть инфекционные агенты (различные бактерии, вирусы), физические факторы (радиоактивное или высокочастотное излучение, повышенная вибрация и др.), химические вещества, заболевания матери, неблагоприятное течение беременности и другие факторы. Например, тяжелые поражения нервной системы, органов слуха, зрения, физического и психического развития возникают при токсоплазмозе, возбудитель которого проникает из организма беременной женщины в плод через плаценту. При резус-конфликте крови матери и плода поражаются подкорковые образования, височные области коры и слуховые нервы. Следствием черепно-мозговых травм в детском возрасте могут быть параличи, расстройства слуха, зрения, нарушения памяти и речи, снижение интеллектуальных возможностей.
Когда повреждение головного мозга, особенно его коры, происходит в раннем возрасте, последствия бывают обычно менее серьезными, чем после аналогичных нарушений у взрослых. Например, у ребенка, уже научившегося говорить, поражение речевой области левого полушария приводит к полной афазии (утрате речи). Однако примерно через год речь восстанавливается. При этом ее центр перемещается в соответствующую зону правого полушария. Такая передача речевого доминирования от левого полушария к правому возможна лишь до 10-летнего возраста. После этого способность к формированию центра речи в любом из полушарий утрачивается.
О громные
возможности образования ассоциативных
связей в нервной системе, отсутствие
узкой специализации нейронов коры
больших полушарий, формирование сложных
«ансамблей нейронов» составляют основу
больших компенсаторных возможностей
коры головного мозга, что используется
в процессе реабилитации лиц с теми или
иными отклонениями в развитии. При
помощи специальных методик дефектолог
может компенсировать нарушенные функции
за счет сохраненных. Например, при
врожденной глухоте или тугоухости
ребенка можно обучить зрительному
восприятию устной речи. В качестве
временного заместителя устной речи
может быть использована дактильная
речь.
громные
возможности образования ассоциативных
связей в нервной системе, отсутствие
узкой специализации нейронов коры
больших полушарий, формирование сложных
«ансамблей нейронов» составляют основу
больших компенсаторных возможностей
коры головного мозга, что используется
в процессе реабилитации лиц с теми или
иными отклонениями в развитии. При
помощи специальных методик дефектолог
может компенсировать нарушенные функции
за счет сохраненных. Например, при
врожденной глухоте или тугоухости
ребенка можно обучить зрительному
восприятию устной речи. В качестве
временного заместителя устной речи
может быть использована дактильная
речь.
1. 3 Строение нейрона. В состав нервной ткани входят высокоспециализированные нервные клетки, называемые нейронами, и клетки нейроглии. Последние тесно связаны с нервными клетками и выполняют опорную, секреторную и защитную функции.
Нейрон – это основная структурная и функциональная единица нервной системы. Нейрон способен генерировать нервные импульсы, проводить возбуждение, принимать, обрабатывать, кодировать, хранить информацию, организовывать реакции на раздражения, устанавливать контакты с другими нейронами, клетками органов и выделять биологически активные вещества. В нем различают тело клетки, или сому, один мало ветвящийся отросток – аксон и много (от 1 до 1000) сильно ветвящихся отростков – дендритов (рис. 1).
Функционально в нейроне выделяют следующие части: воспринимающую – дендриты, мембрана сомы нейрона; интегративную – сома с аксонным холмиком; передающую – аксонный холмик с аксоном.
Тело нейрона в различных отделах нервной системы имеет разную величину (диаметр его колеблется от 4 до 130 мкм) и форму (округлую, уплощенную, многоугольную, овальную). Оно покрыто сложно устроенной мембраной и содержит органоиды, свойственные любой другой клетке: в цитоплазме находятся ядро с одним или несколькими ядрышками, митохондрии, рибосомы, аппарат Гольджи, эндоплазматическая сеть и др.
Характерной особенностью строения нервной клетки является наличие гранулярного ретикулума с большим количеством рибосом (базофильное вещество, вещество Ниссля, тигроидное вещество или тигроид) и нейрофибрилл. С рибосомами в нервных клетках связывают высокий уровень обмена веществ, синтез белка и РНК. Длительное возбуждение нейрона приводит к исчезновению в клетке базофильного вещества, а значит, и к прекращению синтеза специфического белка. У новорожденных нейроны лобной доли коры большого мозга не имеют базофильного вещества. В то же время в структурах, обеспечивающих жизненно важные рефлексы – спинном мозге, мозговом стволе, нейроны содержат большое количество базофильного вещества.
Нейрофибриллы представляют собой тончайшие волоконца, пересекающие тело клетки во всех направлениях и продолжающиеся в отростки. Они принимают участие в хранении и передаче информации.
Тело нейрона (сома), помимо информационной, выполняет трофическую функцию по отношению к своим отросткам и их синапсам. Перерезка аксона или дендрита ведет к гибели отростков, лежащих дистальнее перерезки, а следовательно, и синапсов этих отростков. Сома обеспечивает также рост дендритов и аксона.
Аксон данной клетки имеет постоянный диаметр, в большинстве случаев одет в миелиновую оболочку, образованную из глии, от него отходят отдельные отростки – коллатерали. Аксон имеет разветвленные окончания. В окончаниях находятся митохондрии и секреторные образования. Длина аксона составляет от 0,5 мм до метра и более, его диаметр колеблется от сотых долей микрона (мкм) до 10 мкм. На протяжении первых 50 – 100 мкм от тела клетки аксон не покрыт миелиновой оболочкой. Прилегающий к нему участок тела клетки называют аксонным холмиком.
Аксон передает возбуждение от одного нейрона к другому или рабочему органу. Один аксон может контактировать с 5 тыс. нервных клеток и создавать до 10 тыс. контактов. Обработка поступившей в него информации происходит путем сложных нейрохимических перестроек белковых молекул в нейротрубочках тела нейрона, в результате в аксонном холмике генерируется возбуждение. Участок аксона, не покрытый миелиновой оболочкой, вместе с аксонным холмиком называют начальным сегментом аксона.
По дендритам возбуждение от рецепторов или других нейронов передается к телу клетки. Длина дендрита, как правило, не превышает 300 мкм (хотя длина дендрита афферентного нейрона может достигать метра и более), а диаметр его составляет – 5 мкм. На дендритах имеются боковые отростки (шипики), которые увеличивают их поверхность и являются местами наибольших контактов с другими нейронами. Чем сложнее функция нервной системы, чем больше разных анализаторов посылают информацию к данной структуре, тем больше «шипиков» на дендритах нейронов. Максимальное количество их содержится на пирамидных нейронах двигательной зоны коры большого мозга и достигает нескольких тысяч. За счет «шипиков» воспринимающая поверхность нейрона значительно возрастает и может достигать, например у клеток Пуркинье, 250 000 мкм . Если данный «шипик» или группа «шипиков» длительное время перестает получать информацию, то они исчезают.
Основной особенностью обмена веществ в нейроне является высокая скорость обмена и преобладание аэробных процессов. Потребность мозга в кислороде очень велика. Хотя вес мозга по отношению к весу тела составляет всего 2%, потребление кислорода мозгом достигает в состоянии покоя у взрослых людей 25% от общего его потребления организмом, а у маленьких детей – 50%. Энерготраты мозга составляют 1/6 – 1/8 суточных затрат организма человека.
Все нейроны ЦНС соединяются друг с другом в основном в одном направлении: разветвления аксона одного нейрона контактируют с телом клетки и дендритами другого нейрона.
Нейроны различают по строению и функции. По строению (в зависимости от количества отходящих от тела клетки отростков) различают псевдоуниполярные (оба отростка – аксон и дендрит – выходят из одного полюса клетки), биполярные (с двумя отростками) и мультиполярные (с множеством отростков) нейроны (рис. 2).
П о
функциональным свойствам выделяют
афферентные, чувствительные или
центростремительные нейроны, несущие
возбуждение от рецепторов в ЦНС,
эфферентные, двигательные, мотонейроны,
или центробежные,
передающие возбуждение из ЦНС к
иннервируемому органу, и вставочные,
контактные, или промежуточные нейроны,
соединяющие между собой афферентные и
эфферентные пути.
о
функциональным свойствам выделяют
афферентные, чувствительные или
центростремительные нейроны, несущие
возбуждение от рецепторов в ЦНС,
эфферентные, двигательные, мотонейроны,
или центробежные,
передающие возбуждение из ЦНС к
иннервируемому органу, и вставочные,
контактные, или промежуточные нейроны,
соединяющие между собой афферентные и
эфферентные пути.
Афферентные нейроны относятся к псевдоуниполярным, их тела лежат в спинномозговых ганглиях. Отходящий от тела клетки отросток Т-образно делится на две ветви, одна из которых идет в центральную нервную систему и выполняет функцию аксона, а другая подходит к рецепторам и представляет собой длинный дендрит.
Большинство эфферентных и вставочных нейронов относится к мультиполярным. Мультиполярные вставочные нейроны (самые многочисленные) в большом количестве располагаются в задних рогах спинного мозга, находятся и во всех других отделах ЦНС. Они могут быть и биполярными, как, например, нейроны сетчатки, имеющие короткий ветвящийся дендрит и длинный аксон. Промежуточные нейроны осуществляют связь между эффекторными и рецепторными нейронами.
Мотонейроны располагаются в основном в передних рогах спинного мозга, они передают информацию от нервного центра к исполнительным органам.
С точки зрения выделяемого медиатора нейроны бывают: холинергические, адренергические, дофаминергические, серотонинергические, глицинергические, ГАМК-ергические и т. д. Несмотря на то, что у нейрона множество разветвлений аксона, во всех его окончаниях выделяется один и тот же медиатор (правило Дейла).
По чувствительности к действию раздражителей нейроны делят на моно-, би-, и полисенсорные.
Моносенсорные нейроны располагаются чаще в первичных проекционных зонах коры и реагируют только на сигналы своей сенсорности. Например, значительная часть нейронов первичной зоны зрительной области коры большого мозга реагирует только на световое раздражение сетчатки глаза.
Моносенсорные нейроны подразделяют функционально по их чувствительности к разным качествам одного раздражителя. Так, например, отдельные нейроны слуховой зоны коры большого мозга могут реагировать на предъявления тона в 1000 Гц и не реагировать на тоны другой частоты. Они называются мономодальными. Нейроны, реагирующие на два разных тона, называются бимодальными, на три и более – полимодальными.
Бисенсорные нейроны. Чаще располагаются во вторичных зонах коры какого-либо анализатора и могут реагировать на сигналы как своей, так и другой сенсорности. Например, нейроны вторичной зоны зрительной области коры большого мозга реагируют на зрительные и слуховые раздражения.
Полисенсорные нейроны. Это чаще всего нейроны ассоциативных зон мозга; они способны реагировать на раздражение слуховой, зрительной, кожной и других рецептивных систем.
1. 4 Нейроглия. Нейроглия – это вспомогательная и очень важная составная часть нервной ткани, связанная с нейронами генетически, морфологически и функционально. Эти клетки заполняют в мозге все пространство между нейронами, они мельче нейронов в 3 – 4 раза. Каждый нейрон окружен несколькими клетками нейроглии, которые равномерно распределены по всему мозгу и составляют около 40% его объёма. В коре больших полушарий их примерно в 5 раз больше, чем нервных клеток.
Нейроглия выполняет опорную, трофическую, защитную, изоляционную функции. Клетки нейроглии, возможно, принимают участие в обмене информацией внутри нейронного ансамбля, а также в условно-рефлекторной деятельности мозга и в процессах памяти. Глиальные клетки обладают способностью перемещаться в пространстве по направлению к наиболее активным нейронам. Это наблюдается при различных афферентных раздражениях и при мышечной нагрузке. Например, уже через 20 мин. плавания у крыс было обнаружено увеличение числа глиальных клеток вокруг мотонейронов переднего рога спинного мозга.
С возрастом количество нейронов в ЦНС уменьшается, а клеток нейроглии – увеличивается, так как последние, в отличие от нейронов, сохраняют способность к делению.
Нейроглия по своему происхождению подразделяется на макроглию и микроглию. Макроглия, как и нейроны, возникает из эктодермы, а микроглия развивается из мезодермы и является производным мезенхимы. В состав макроглии входят эпендима, астроглия и олигодендроглия.
Клетки эпендимы называются эпендимоцитами. Она состоит из клеток цилиндрической формы, выстилающих желудочки головного мозга и центральный канал спинного мозга. Играет роль барьера между спинномозговой жидкостью и тканью мозга; выполняет, по-видимому, и секреторную функцию.
А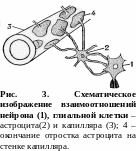 строглия
(около
60% от общего числа клеток нейроглии)
состоит из клеток – астроцитов, названных
так потому, что от них радиально отходят
многочисленные отростки. Астроглия
выполняет опорную и трофическую функции
(рис. 3). Отростки астроцитов образуют
также пограничный слой на поверхности
головного и спинного мозга – пограничную
мембрану мягкой мозговой оболочки,
прилегающую к базальной мембране. Это
основной элемент гемато-энцефалического
барьера; регулирует водно-солевой обмен
нервной ткани. Астроциты образуют
мостики между капиллярами и эпендимой,
выстилающей полости желудочков мозга.
Таким образом обеспечивается обмен
между кровью и цереброспинальной
жидкостью, т. е. астроциты выполняют
транспортную функцию.
строглия
(около
60% от общего числа клеток нейроглии)
состоит из клеток – астроцитов, названных
так потому, что от них радиально отходят
многочисленные отростки. Астроглия
выполняет опорную и трофическую функции
(рис. 3). Отростки астроцитов образуют
также пограничный слой на поверхности
головного и спинного мозга – пограничную
мембрану мягкой мозговой оболочки,
прилегающую к базальной мембране. Это
основной элемент гемато-энцефалического
барьера; регулирует водно-солевой обмен
нервной ткани. Астроциты образуют
мостики между капиллярами и эпендимой,
выстилающей полости желудочков мозга.
Таким образом обеспечивается обмен
между кровью и цереброспинальной
жидкостью, т. е. астроциты выполняют
транспортную функцию.
О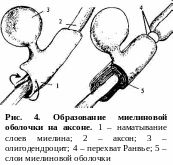 лигоденроглия
(около 25—30%) состоит из клеток, называемых
олигодендроцитами, имеющих малое
количество отростков. Отличаются высоким
уровнем белкового и нуклеинового обмена;
ответственны за транспорт веществ в
нейроны. Они находятся в сером и белом
веществе головного и спинного мозга, а
также за пределами центральной нервной
системы. В сером веществе центральных
отделов нервной системы они окружают
тела нейронов и их отростков, в белом
веществе располагаются рядами или
группами, а за пределами центральной
нервной системы сопровождают отростки
нервных клеток и известны под названием
шванновских клеток, или нейролеммоцитов.
Олигодендроциты синтезируют миелин –
вещество белого цвета липоидной природы,
которое обладает хорошими изоляционными
свойствами (рис. 4). Изолируя отростки
нервных клеток, олигоденроциты
препятствуют рассеиванию нервного
возбуждения. Кроме того, олигодендроциты
принимают участие в питании нейронов
и водном обмене мозга, поскольку всегда
контактируют с кровеносными сосудами
и, по некоторым данным, вместе с астроцитами
участвуют в хранении следов возбуждения
в нервной системе.
лигоденроглия
(около 25—30%) состоит из клеток, называемых
олигодендроцитами, имеющих малое
количество отростков. Отличаются высоким
уровнем белкового и нуклеинового обмена;
ответственны за транспорт веществ в
нейроны. Они находятся в сером и белом
веществе головного и спинного мозга, а
также за пределами центральной нервной
системы. В сером веществе центральных
отделов нервной системы они окружают
тела нейронов и их отростков, в белом
веществе располагаются рядами или
группами, а за пределами центральной
нервной системы сопровождают отростки
нервных клеток и известны под названием
шванновских клеток, или нейролеммоцитов.
Олигодендроциты синтезируют миелин –
вещество белого цвета липоидной природы,
которое обладает хорошими изоляционными
свойствами (рис. 4). Изолируя отростки
нервных клеток, олигоденроциты
препятствуют рассеиванию нервного
возбуждения. Кроме того, олигодендроциты
принимают участие в питании нейронов
и водном обмене мозга, поскольку всегда
контактируют с кровеносными сосудами
и, по некоторым данным, вместе с астроцитами
участвуют в хранении следов возбуждения
в нервной системе.
Некоторые клетки нейроглии выделяют вещества, влияющие на состояние возбудимости нервных клеток. Отмечено, что при разных психических состояниях изменяется секреция этих клеток.
Микроглия представлена клетками микроглии, или клетками Гортега, которые имеют небольшие древовидные отростки и отличаются малыми размерами. В нормальном состоянии они располагаются поодиночке в различных отделах центральной нервной системы, при раздражении могут изменять свою форму, округляются и приобретают способность к амебоидному движению и фагоцитозу остатков погибших нервных клеток и посторонних частиц. Кроме того, клетки микроглии запасают жир.
1. 5 Нервные волокна, их виды, строение, свойства, возрастные изменения в связи с миелинизацией. Отросток нервной клетки, покрытый оболочками из глиальных клеток, называется нервным волокном. Центральную часть любого отростка нервной клетки (аксона или дендрита) называют осевым цилиндром. Осевой цилиндр покрыт оболочкой – аксолеммой, под которой находится аксоплазма, содержащая нейрофибриллы и большое количество митохондрий.
В различных отделах нервной системы нервные волокна значительно различаются по своему строению, в связи с чем делятся на два типа: безмиелиновые и миелиновые. Миелиновая оболочка образуется шванновскими клетками путем их многократного накручивания вокруг осевого цилиндра и располагается не сплошной линией, а сегментами длиной 0,5 – 2 мм. Пространство между сегментами (1 – 2 мкм) называют перехватом Ранвье. В области перехватов Ранвье миелиновая оболочка отсутствует, и концы шванновских клеток плотно прилегают к аксолемме (рис. 4). Наружная мембрана шванновских клеток, покрывающая миелин, образует самую верхнюю оболочку нервного волокна, которую называют шванновской оболочкой, или неврилеммой. Безмиелиновые волокна отличаются тем, что в них не развивается миелиновая облочка и их осевые цилиндры покрыты только шванновскими клетками (шванновской оболочкой).
К миелинизированным относятся волокна соматической нервной системы и некоторые волокна вегетативной нервной системы. К безмиелиновым относится большинство волокон вегетативной нервной системы.
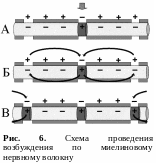
В безмякотных нервных волокнах возбуждение распространяется непрерывно вдоль всей мембраны, от возбужденного участка к невозбужденному, расположенному рядом, и сопровождается бóльшим расходом энергии, по сравнению с миелиновым волокном (рис. 5). Скорость проведения возбуждения относительно невелика. В отличие от этого в миелиновых волокнах возбуждение может распространяться только скачкообразно, «перепрыгивая» по перехватам Ранвье через участки волокна, покрытые миелином (рис. 6). Такое проведение называется сальтаторным. Возбуждение по мякотному нервному волокну проводится с очень большой скоростью. Расход энергии при этом невелик. Нервный импульс, возникший в одном перехвате Ранвье, способен вызывать возбуждение не только в лежащем рядом, но и в соседних 2–3 перехватах. Это создает гарантию проведения возбуждения по волокну, даже если 1 – 2 лежащих рядом перехвата повреждены.
Н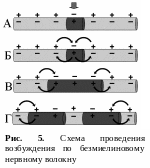
 ервные
волокна делят на три группы, учитывая
их строение и скорость проведения
возбуждения: А, В и С. В группу «А» входят
толстые миелиновые волокна, с диаметром
15 мкм и скоростью проведения возбуждения
от 10 до 150 м/с. В группу «В» – тонкие
миелиновые волокна диаметром 10 мкм,
скоростью проведения возбуждения до
10 м/с; в группу «С» – тонкие безмиелиновые,
диаметр 5 мкм, скорость поведения
возбуждения до 1 м/с. В волокнах любого
типа возбуждение распространяется без
затухания (без декремента).
ервные
волокна делят на три группы, учитывая
их строение и скорость проведения
возбуждения: А, В и С. В группу «А» входят
толстые миелиновые волокна, с диаметром
15 мкм и скоростью проведения возбуждения
от 10 до 150 м/с. В группу «В» – тонкие
миелиновые волокна диаметром 10 мкм,
скоростью проведения возбуждения до
10 м/с; в группу «С» – тонкие безмиелиновые,
диаметр 5 мкм, скорость поведения
возбуждения до 1 м/с. В волокнах любого
типа возбуждение распространяется без
затухания (без декремента).
Основные свойства нервных волокон заключаются в следующем: высокая возбудимость и лабильность, низкий уровень обмена веществ, относительная неутомляемость, большая скорость проведения возбуждения. Проведение возбуждения по нервам подчиняется ряду законов: физиологической и анатомической целостности нерва, двустороннего проведения возбуждения от места возникновения, изолированного проведения возбуждения. Нервные импульсы распространяются по нервным волокнам без затухания или без декремента.
Миелинизация нервных волокон осуществляется в центробежном направлении, отступя несколько микрон от тела клетки к периферии нервного волокна. Отсутствие миелиновой оболочки резко ограничивает функциональные возможности нервного волокна, обусловливает диффузные, слабо координированные реакции.
По мере развития миелиновой оболочки возбудимость нервных волокон повышается. Раньше других начинают миелинизироваться периферические нервы, затем волокна спинного мозга, стволовой части головного мозга, мозжечка и позже – больших полушарий головного мозга. Этот процесс начинается на 4-м месяце внутриутробного периода и завершается в основном к 3 годам. Известно, что у новорожденного ребенка миелинизировано примерно 2/3 волокон головного мозга. Примерно к 12 годам завершается следующий этап миелинизации. Это соответствует тому, что у ребенка уже формируется функция внимания, он достаточно хорошо владеет собой. Вместе с тем полностью процесс миелинизации заканчивается только при завершении полового созревания. Таким образом, процесс миелинизации является показателем созревания ряда психических функций.
1. 6 Строение синапса. Синапс – это морфофункциональное образование, которое обеспечивает передачу сигнала с нейрона на другой нейрон или с нейрона на эффекторную клетку (от греческого слова «синапто» – контактировать). Термин и понятие «синапс», «синаптическая передача» был введен в физиологию Ч. Шеррингтоном в 1897 г.
Чем больше синапсов на нервной клетке, тем больше воспринимается различных раздражений и, следовательно, шире сфера влияний на ее деятельность и возможность участия нервной клетки в разнообразных реакциях организма. На одном крупном мотонейроне насчитывают до 20 000 синапсов.
Количество синаптических контактов неодинаково на теле и отростках нейрона и очень вариабельно в различных отделах центральной нервной системы. Тело нейрона в среднем на 38% покрыто синапсами. Наибольшее число (до 50%) синапсов находится на дендритах. Особенно густо они покрывают средние части и окончания дендритных отростков, причем многие контакты расположены на специальных шипиках, которые еще больше увеличивают воспринимающую поверхность нейрона. Чем сложнее интегративная функция нейрона, тем большее развитие имеют аксодендритические синапсы (в первую очередь те, которые расположены на шипиках). Особенно они характерны для нейрональных связей пирамидных клеток в коре больших полушарий.
По способу передачи возбуждения синапсы подразделяют на электрические и химические и смешанные.
По местоположению синапсы подразделяются на нейро-мышечные (аксон нейрона контактирует с мышечной клеткой); нейро-секреторные (аксон нейрона контактирует с секреторной клеткой); нейро-нейрональные (аксон нейрона контактирует с другим нейроном)
По физиологическому эффекту синапсы подразделяют на возбуждающие и тормозные.
Синапс состоит из пресинаптического и постсинаптического отделов, между которыми имеется небольшое пространство, получившее название синаптической щели.
У электрического синапса клеточные мембраны соседних нейронов тесно прилегают друг к другу, так что между ними остается только очень узкая щель шириной 2 – 5 нм. Обе мембраны соединяются посредством поперечных каналов, образованных особыми белками-коннексонами. Эти каналы не сообщаются с внеклеточным пространством и отсутствуют в других участках мембраны. Они имеют низкую утечку во внешнюю среду и передают нервные импульсы с использованием того же механизма, что и при передаче по нервному волокну.
Электрические синапсы находятся там, где нужна быстрая передача, они биполярны, симметричны, сравнительно мало утомляемы, устойчивы к изменениям внешней и внутренней среды, проводят только возбуждение и возбуждением могут охватывать сразу несколько нейронов. У человека, также как и у млекопитающих, электрические синапсы находятся в нескольких областях мозга: в мезэнцефальном ядре тройничного нерва, в вестибулярном ядре Дейтерса, в нижней оливе, между некоторыми клетками в сетчатке.
Химические синапсы очень специфические, не симметричные, односторонние, между мембранами имеется щель. На прохождение щели уходит время. В отличие от электрических проводят как возбуждение, так и торможение. Химические синапсы обладают высокой утомляемостью. Это объясняется истощением запасов медиатора.
С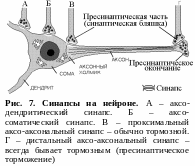 инапсы,
образованные аксоном и телом (сомой)
клетки, называют аксо-соматическими,
аксоном и дендритом – аксо-дендритическими,
контакты между аксонами двух нейронов
получили название аксо-аксональных
синапсов (рис. 7).
инапсы,
образованные аксоном и телом (сомой)
клетки, называют аксо-соматическими,
аксоном и дендритом – аксо-дендритическими,
контакты между аксонами двух нейронов
получили название аксо-аксональных
синапсов (рис. 7).
Пресинаптический отдел химического синапса представлен конечной веточкой аксона, которая на расстоянии 200 – 300 мкм от контакта теряет миелиновую оболочку и на конце утолщается (рис. 8). Это утолщение называется синаптической бляшкой. В пресинаптическом отделе синапса содержится большое количество митохондрий и пузырьков (везикул) округлой или овальной формы размером от 0,02 до 0,05 мкм. В везикулах содержится вещество, обеспечивающее передачу возбуждения с одного нейрона на другой, которое называют медиатором. Везикулы концентрируются вдоль поверхности пресинаптической мембраны, находящейся против синаптической щели, ширина которой равна 0,0012 – 0,03 мкм.
Постсинаптический отдел синапса образуется мембраной сомы нейрона или его отростков, а в концевой пластинке (синапс между окончанием аксона и мышцей) – мембраной мышечного волокна. Постсинаптическая мембрана несет на себе белковые хеморецепторы к биологически активным веществам. Важная особенность рецепторов постсинаптической мембраны – их химическая специфичность, т. е. способность вступать в биохимическое взаимодействие только с определенным видом медиатора.
П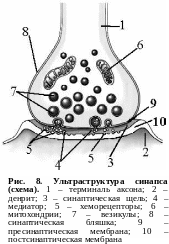 ресинаптическая
и постсинаптичес-кая мембраны имеют
специфические особенности строения,
связанные с передачей возбуждения: они
несколько утолщены (их диаметр около
0,005 мкм).
Длина этих участков составляет
150 – 450 мкм.
Утолщения могут быть сплошными и
прерывистыми. Постсинаптическая мембрана
у некоторых синапсов может быть
складчатой, что увеличивает поверхность
соприкосновения ее с медиатором.
ресинаптическая
и постсинаптичес-кая мембраны имеют
специфические особенности строения,
связанные с передачей возбуждения: они
несколько утолщены (их диаметр около
0,005 мкм).
Длина этих участков составляет
150 – 450 мкм.
Утолщения могут быть сплошными и
прерывистыми. Постсинаптическая мембрана
у некоторых синапсов может быть
складчатой, что увеличивает поверхность
соприкосновения ее с медиатором.
Проходящий по терминали аксона нервный импульс достигает пресинаптической мембраны, что приводит к её деполяризации и открытию кальциевых пор (каналов). Кальций мигрирует через эти каналы внутрь аксонной терминали и способствует слиянию везикул с пресинаптической мембраной и опорожнению медиатора (экзоцитозу) в синаптическую щель. Количество медитора, которое освобождается из одного синаптического пузырька, называется квантом медиатора. Под действием одного нервного импульса в синаптическую щель освобождается 100 – 200 квантов медиатора.
Выделившийся медиатор диффундирует к постсинаптической мембране. Этот процесс требует около 0,5 мс и вносит существенный вклад в задержку передачи. Взаимодействие медиатора со своими рецепторами вызывает возбуждение или торможение нейрона, сокращение мышечной клетки, образование и выделение гормонов секреторными клетками. Передача возбуждения происходит только в одном направлении: от синаптического окончания к постсинаптической мембране.
В синапсах ЦНС, как и в концевой пластинке, в состоянии относительного покоя отмечено спонтанное разрушение везикул и выход квантов медиатора в синаптическую щель. Этого недостаточно для возникновения возбуждения в постсинаптической мембране, однако синапс поддерживается в состоянии готовности к передаче.
Инактивация (полная потеря активности) медиатора осуществляется путем гидролитического расщепления с помощью соответствующих ферментов. Ацетилхолин (АХ) расщепляется ферментом холинэстеразой, норадреналин (НА) и адреналин – моноаминоксидазой (МАО) и катехолоксиметилтрансферазой (КОМТ). Продукты расщепления медиатора снова поступают в кровь и циркулируют, как его предшественники.
Другой путь удаления медиатора из синаптической щели – «обратный захват» его пресинаптическими окончаниями (пиноцитоз) и обратный аксонный транспорт, особенно выраженный для катехоламинов. Это создает возможность их быстрого поступления в синаптическую щель под влиянием нового нервного импульса.
Для обеспечения нормальной работы химического синапса требуется поддержание необходимого количества везикул и медиатора. Некоторые медиаторы (например, ацетилхолин) синтезируются в цитоплазме синаптического окончания, и там же молекулы медиатора депонируются в синаптических пузырьках. Ферменты, необходимые для синтеза медиатора, образуются в теле нейрона и доставляются в синаптическое окончание путем медленного (1–3 мм/сут) аксонного транспорта. Другие медиаторы (пептиды и др.) синтезируются и упаковываются в везикулы в теле нейрона, готовые синаптические пузырьки доставляются в синаптичекую бляшку за счет быстрого (400 мм/сут) аксонного транспорта. Синтез медиатора и образование синаптических пузырьков осуществляется непрерывно.
В ЦНС выделяют два типа нейронов: возбуждающие и тормозящие. Возбуждающие нейроны выделяют в своих синапсах такие медиаторы, как ацетилхолин (АХ), норадреналин, серотонин, кислые аминокислоты. Тормозящие нейроны вызывают торможение за счет медиаторов, таких как глицин, β-аланин, γ-аминомасляниая кислота (ГАМК), дофамин. В то же время установлено, что один и тот же медиатор, например АХ, может выделяться как возбуждающими, так и тормозными синапсами, и, соответственно, вызывать как возбуждение, так и торможение. Поэтому можно говорить об амбивалентности медиаторов.
Каждый синаптический контакт на теле или отростках нейрона способствует развитию местных процессов возбуждения или торможения. На мембране нейрона происходит их алгебраическое суммирование. При одновременной активации нескольких возбуждающих синапсов их общий эффект представляет собой сумму отдельных местных (локальных) процессов возбуждения. При одновременном возникновении двух различных синаптических влияний (возбуждающих и тормозящих) происходит взаимное вычитание их эффектов. В конечном итоге реакция нервной клетки определяется суммой всех синаптических влияний.
Активная деятельность нервных клеток способствует улучшению синаптического взаимодействия между ними: росту площади синаптических контактов, ускорению проведения возбуждения через синапсы и т. д. Это наблюдается, например, при двигательной деятельности. Как показали эксперименты на животных, ежедневная принудительная ходьба их по 1 часу приводит к значительной активации синапсов в соответствующих рефлекторных дугах спинного мозга.
Синапсы, являются самым чувствительным местом нервной системы. Именно синаптические контакты между нейронами первыми реагируют на различные повреждающие воздействия. При травмах нервной ткани, отеках, нарушениях кровообращения, рентгеновских облучениях, отравлениях и других неблагоприятных воздействиях в синапсах сначала возникают обратимые изменения - сильное набухание (увеличение их размеров в несколько раз) и деформация, а затем происходят необратимые изменения синапсов - их разрушение, в результате чего полностью нарушается нервная деятельность.
1. 7 Рефлекс и рефлекторная дуга. Ответную реакцию организма на раздражение рецепторов, осуществляющуюся с участием центральной нервной системы, называют рефлексом. Раздражителем, вызывающим рефлекторную реакцию, может быть любое изменение внешней или внутренней среды организма.
Время от момента нанесения раздражения до появления ответной реакции называют временем рефлекса. Время рефлекса зависит от количества нейронов (синапсов), входящих в рефлекторную дугу, а также от интенсивности раздражения: чем больше сила раздражения, тем меньше время рефлекса.
Основным механизмом деятельности ЦНС является рефлекс. Впервые понятие рефлекса как отражательной деятельности было введено Рене Декартом в начале 17 века. Это представление было развито чешским физиологом Г. Прохаской (18 в.). Узловым моментом развития рефлекторной теории считают классический труд И. М. Сеченова (1863) «Рефлексы головного мозга», в котором впервые провозглашен тезис о том, что все виды сознательной и бессознательной жизни человека представляют собой рефлекторные реакции.
В основе рефлекторной теории Сеченова лежат четыре принципа:
структурности (структурной основой рефлекса является рефлекторная дуга);
детерминизма (принцип причинно-следственных отношений) – ни одна ответная реакция организма не бывает без причины – воздействия соответствующего раздражителя;
анализа и синтеза (любое воздействие на организм сначала анализируется, затем обобщается, т. е. вырабатывается ответная реакция);
обратной связи (отображающий точность реакций и адаптацию), особенно развитый в трудах академика П. К. Анохина.
Структурной основой рефлекса является рефлекторная дуга. Рефлекторной дугой называют путь, по которому нервный импульс распространяется от рецептора к эффектору. Функционально она состоит из афферентного, центрального и эфферентного звеньев, связанных между собой синапсами. Морфологически в рефлекторной дуге выделяют 5 отделов:
1) рецептор (воспринимает раздражение из внешней или внутренней среды и преобразует его энергию в энергию нервного импульса);
2) чувствительное (афферентное) волокно, по которому возбуждение передается в ЦНС;
3) ЦНС (нервный центр), где происходит анализ поступивших сигналов и выработка ответной реакции;
4) эфферентное нервное волокно, по которому возбуждение направляется из ЦНС к рабочему органу.
5) эффектор или рабочий орган (мышца или железа).
В
зависимости от сложности структуры
рефлекторной дуги различают моно- и
полисинаптические рефлексы. В простейшем
случае импульсы, поступающие в ЦНС по
афферентным путям, переключаются
непосредственно с афферентного нейрона
на эфферентный, т. е. в системе рефлекторной
дуги имеется одно синаптическое
соединение (рис. 9А). Такая рефлекторная
дуга называется моносинаптической или
двухнейронной (например, сухожильные
рефлексы: ахиллов, коленный и др.).
Особенностью
двухнейронной дуги является то, что и
рецептор и эффектор рефлексов,
осуществляющихся по э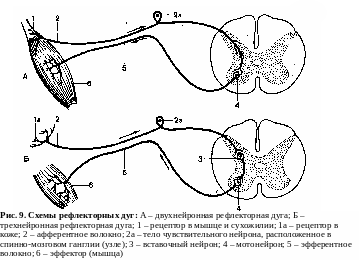 тому
типу, лежат в одном и том же органе.
Моносинаптических
рефлексов в ЦНС сравнительно немного
– около 1 %.
тому
типу, лежат в одном и том же органе.
Моносинаптических
рефлексов в ЦНС сравнительно немного
– около 1 %.
Наличие в структуре рефлекторной дуги двух и более синаптических переключений (т. е. трех и более нейронов) позволяет характеризовать ее как полисинаптическую. Полисинаптическая рефлекторная дуга помимо афферентного и эфферентного нейронов содержит еще 1 и более вставочных нейронов (рис. 9Б).
Дендриты афферентных нейронов начинаются от рецепторов и образуют афферентные нервные волокна. Тела афферентных нейронов, как правило, лежат в спинномозговых ганглиях вне ЦНС. Их аксоны в составе задних корешков входят в спинной мозг. Вставочные нейроны располагаются в разных отделах спинного и головного мозга. Их количество может сильно варьировать. Тела эфферентных нейронов (двигательных) нейронов находятся в передних рогах спинного мозга. Их аксоны в составе передних корешков спинного мозга выходят их ЦНС и образуют эфферентные нервные волокна.
Рефлекторный акт не заканчивается ответной реакцией организма на раздражение. В работающем органе раздражаются рецепторы, и возникающее в них возбуждение по афферентным волокнам возвращается в центральную нервную систему. Каждый орган, будь это скелетные мышцы, осуществляющие движение, или ритмически сокращающаяся сердечная мышца, или мышцы постоянно пульсирующих сосудов, дыхательные мышцы и т. д., благодаря обратной импульсации «сообщает» нервным центрам о своем состоянии. В ответ на полученный афферентный сигнал нервный центр в каждый данный момент вносит свои поправки в осуществляющийся рефлекторный акт. Поэтому принято говорить не о рефлекторной дуге, а о рефлекторном кольце.
1. 8 Классификация рефлексов. Классифицируя многообразные рефлексы организма человека и животного, учитывают их различные признаки и проявления.
Все рефлексы по происхождению делят на безусловные – врожденные – видовые рефлексы и условные – приобретенные в процессе индивидуальной жизни животного или человека, выработанные при определенных условиях.
На основании биологического значения тех или иных рефлексов для организма их делят на оборонительные, или защитные, направленные на удаление от раздражителя; пищевые, обеспечивающие добывание, потребление и переваривание пищи; половые, связанные с продолжением рода; ориентировочные, или исследовательские, проявляющиеся в повороте тела и движении в сторону нового раздражителя; позно-тонические, или рефлексы положения тела в пространстве, и локомоторные – перемещения тела в пространстве.
В зависимости от расположения рецепторов рефлекторной дуги, различают экстероцептивные рефлексы, возникающие в ответ на раздражение рецепторов поверхности тела, проприоцептивные – на раздражение рецепторов мышц, сухожилий и суставов, висцероцептивные – на раздражение рецепторов внутренних органов.
Классифицируют рефлексы и по органам, отвечающим на раздражение. При этом говорят о сердечных, дыхательных, сосудистых и других рефлексах.
Рефлексы различают также по характеру ответных реакций: секреторные, выражающиеся в выделении секрета, выработанного железой, трофические, связанные с изменением обмена веществ, и двигательные, или моторные, характеризующиеся сократительной деятельностью поперечнополосатых и гладких мышц. Последние составляют самую разнообразную группу рефлексов. При раздражении кожи возникают сгибательные, потирательные рефлексы, рефлексы почесывания и др. Раздражением кожи губ у ребенка вызывается сосательный рефлекс, роговицы глаза – мигательный. Сужение зрачка при действии света и его расширение в темноте представляют собой зрачковый рефлекс.
Двигательные проприоцептивные рефлексы возникают при раздражении рецепторов мышц и сухожилий. Так, при ударе по сухожилию четырехглавой мышцы бедра в результате ее растяжения возникает рефлекторное разгибание ноги в колене – коленный рефлекс, при ударе по ахиллову сухожилию – ахиллов рефлекс.
Рефлексы, заключающиеся в сужении и расширении сосудов, называются сосудодвигательными.
Двигательные рефлексы, возникающие при раздражении рецепторов гладкой мускулатуры внутренних органов, носят название висцеромоторных. С ними связаны движения желудка, кишечника, мочевого пузыря, мочеточников и др.
Все описанные выше рефлексы в зависимости от того, какие отделы центральной нервной системы участвуют в их осуществлении, делят на спинальные (осуществляющиеся с участием нейронов спинного мозга), бульбарные (с участием нейронов продолговатого мозга), мезенцефальные (с участием среднего мозга), диэнцефальные (с участием промежуточного мозга) и кортикальные (с участием нейронов коры больших полушарий головного мозга).
Примером спинальных рефлексов могут быть сгибательный, возникающий при пощипывании лапки лягушки пинцетом, потирательный – при раздражении кожи лягушки бумажкой, смоченной серной кислотой, и др. К ним же относятся рефлексы сухожильного типа.
Сосательный, мигательный рефлексы осуществляются с участием продолговатого, зрачковый – среднего мозга. Регуляция любой функции осуществляется с участием разных отделов центральной нервной системы. Поэтому классификация рефлексов по отделам мозга, участвующим в их осуществлении, относительна. Речь идет только о преимущественном значении нейронов того или иного отдела центральной нервной системы.
1. 9 Классификация и строение рецепторов. Живые организмы не могут существовать, не получая информации из внешней и внутренней среды. Восприятие энергии раздражителя и преобразование ее в нервный импульс осуществляют рецепторы.
Рецептор представляет собой окончание дендрита афферентного аксона или комплекс нервной и эпителиальной клеток, специально приспособленный для восприятия определенного типа раздражителей. Рецепторы являются начальными звеньями любой рефлекторной дуги, а также участвуют в оценке параметров полезного приспособительного результата в функциональных системах организма.
Дендрит каждого афферентного нейрона выходит на периферию и разветвляется на многочисленные тончайшие веточки – терминали. На конце каждой терминали находится утолщение – рецепторная бляшка.
По строению рецепторы подразделяют на первичные (первичночувствующие) и вторичные (вторичночувствующие) (рис. 10).
К первичным относят такие рецепторы, у которых действие раздражителя воспринимается непосредственно рецепторной бляшкой, которая может быть либо свободной, т. е. не иметь дополнительных образований, либо инкапсулированной. В этом случае рецепторная бляшка заключена в особую капсулу, осуществляющую первичное преобразование энергии раздражителя. К первичночувствующим рецепторам относятся: обонятельные, тактильные рецепторы и мышечные веретена.
К вторичным
относят
такие рецепторы, у которых действие
раздражителя воспринимается
специализированной рецептирующей
клеткой не нервного происхождения.
Возбуждение, возникшее в рецептирующей
клетке, передается через синапс на
чувствительный нейрон, который
возбуждается не непосредственно
стимулом, а опосредовано (вторично)
потенциалом рецептирующей клетки. К
вторичночувствующим рецепторам
относятся: рецепторы слуха, зрения,
вкуса, вестибулярные рецепторы.
вторичным
относят
такие рецепторы, у которых действие
раздражителя воспринимается
специализированной рецептирующей
клеткой не нервного происхождения.
Возбуждение, возникшее в рецептирующей
клетке, передается через синапс на
чувствительный нейрон, который
возбуждается не непосредственно
стимулом, а опосредовано (вторично)
потенциалом рецептирующей клетки. К
вторичночувствующим рецепторам
относятся: рецепторы слуха, зрения,
вкуса, вестибулярные рецепторы.
По расположению рецепторы подразделяют на: экстерорецепторы – воспринимают раздражители из внешней среды организма; интерорецепторы – воспринимают раздражители из внутренней среды организма. Они в свою очередь подразделяются на проприорецепторы – специализированные рецепторы опорно-двигательной системы (мышц, сухожилий, суставов), вестибулорецепторы – воспринимают положение тела в пространстве и висцерорецепторы – получают сигналы от внутренних органов.
По разнообразию воспринимаемых раздражителей рецепторы подразделяют на: мономодальные – приспособленные для восприятия только одного вида раздражителя, и полимодальные – приспособленные для восприятия различных раздражителей.
По модальности рецепторы подразделяют на: хеморецепторы – воспринимают действие химических веществ; фоторецепторы – воспринимают световые раздражители; механорецепторы – воспринимают давление, вибрацию, перемещение, степень растяжения; терморецепторы – чувствительны к изменениям температуры; ноцицепторы – воспринимают болевое раздражение.
По способности воспринимать раздражитель, находящийся на расстоянии от рецептора или при непосредственном контакте с ним, различают дискантные и контактные рецепторы
Все рецепторы обладают следующими свойствами.
1. Специфичность – способность воспринимать определенный, т. е. адекватный данному рецептору, раздражитель. Эта способность рецепторов сформировалась в процессе эволюции.
2. Высокая чувствительность – способность реагировать на очень малые по интенсивности параметры адекватного раздражителя. Например, для возбуждения фоторецепторов сетчатки глаза достаточно нескольких, а иногда и одного, квантов света. Обонятельные рецепторы информируют организм о появлении в атмосфере единичных молекул пахучих веществ.
3. Способность к ритмической генерации импульсов возбуждения в ответ на однократное действие раздражителя.
4. Возбуждение рецепторов наступает после некоторого латентного периода и имеет последействие.
5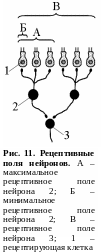 .
Способность к адаптации – т. е. способность
приспосабливаться ("привыкать") к
постоянно действующему стимулу. Адаптация
может выражаться в снижении активности
рецептора и частоты генерации импульсов
возбуждения, вплоть до полного его
прекращения. В зависимости от скорости
адаптации различают: быстроадаптирующиеся
(тактильные), медленноадаптирующиеся
(терморецепторы), неадаптирующиеся
(болевые, вестибулярные и проприорецепторы).
.
Способность к адаптации – т. е. способность
приспосабливаться ("привыкать") к
постоянно действующему стимулу. Адаптация
может выражаться в снижении активности
рецептора и частоты генерации импульсов
возбуждения, вплоть до полного его
прекращения. В зависимости от скорости
адаптации различают: быстроадаптирующиеся
(тактильные), медленноадаптирующиеся
(терморецепторы), неадаптирующиеся
(болевые, вестибулярные и проприорецепторы).
Выделяют несколько видов адаптации: изменение возбудимости рецептора в сторону снижения – десенсибилизация; изменение возбудимости в сторону повышения – сенсибилизация.
При действии сильных раздражителей, порог чувствительности повышается. Например: если человек вышел из тепла сразу в холод – то человеку вначале холодно, но затем рецепторы повышают свой порог и человеку становится тепло.
Р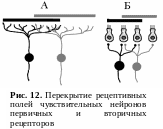 ецептивное
поле.
Рецептивным
полем нейрона
называют множество рецепторов,
функционально связанных с этим нейроном.
Рецептивное поле нейрона представляет
собой динамическое образование – один
и тот же нейрон в различные моменты
времени может оказаться функционально
связанным с различным количеством
рецепторов. Максимальная величина
рецептивного поля какого-либо нейрона
соответствует количеству рецепторов,
которые связаны с эти нейроном
морфологически, а минимальная величина
может ограничиваться всего одним
рецептором (рис. 11).
ецептивное
поле.
Рецептивным
полем нейрона
называют множество рецепторов,
функционально связанных с этим нейроном.
Рецептивное поле нейрона представляет
собой динамическое образование – один
и тот же нейрон в различные моменты
времени может оказаться функционально
связанным с различным количеством
рецепторов. Максимальная величина
рецептивного поля какого-либо нейрона
соответствует количеству рецепторов,
которые связаны с эти нейроном
морфологически, а минимальная величина
может ограничиваться всего одним
рецептором (рис. 11).
У первичных рецепторов зоны ветвления периферических отростков чувствительных нейронов могут перекрывать друг друга (рис. 12, А). У вторичных рецепторов одна рецептирующая клетка может контактировать с несколькими чувствительными нейронами, т. е. может входить в состав рецептивных полей различных нейронов (рис. 12, Б).
Рецептивным полем рефлекса называют совокупность рецепторов, раздражение которых вызывает данный тип рефлекса.
1. 10 Понятие о нервном центре. Свойства нервных центров. В сложных многоклеточных организмах животных и человека отдельная нервная клетка не в состоянии регулировать какие-либо функции. Все основные формы деятельности нервной системы связаны с участием в ее функциях определенных групп нервных клеток – нервных центров. Нервный центр – центральный (третий) компонент рефлекторной дуги, где происходит переработка информации, вырабатывается программа действия, формируется эталон результата.
Нервным центром называется совокупность нейронов, расположенных на различных уровнях ЦНС и регулирующих сложный рефлекторный процесс или функцию. В нервном центре выделяют следующие отделы: низший или исполнительный, рабочий и высший или регуляторный.
Низший (исполнительный) отдел нервного центра локализован в спинном мозге и передает информацию от рабочего отдела к рабочим органам.
Рабочий отдел нервного центра – это отдел, ответственный за осуществление данной функции, как правило, находится в мозговом стволе.
Высший (регуляторный) отдел нервного центра расположен в коре больших полушарий мозга и регулирует активность рабочего отдела нервного центра; вмешивается в регуляцию функций эпизодически, т. е. в тех случаях, когда необходимо внести поправку в автоматическую деятельность рабочего отдела. Высшие отделы вмешиваются в работу нервного центра по условно-рефлекторному механизму.
Активность регуляторного (высшего) отдела нервного центра зависит от состояния рабочего отдела, который получает афферентную информацию и от внешних стимулов среды.
Вышеизложенное можно рассмотреть на примере дыхательного центра. Исполнительный (низший) отдел дыхательного нервного центра расположен в передних рогах грудного отдела спинного мозга и транслирует приказы рабочего центра к дыхательным мышцам. Рабочий отдел представлен центрами вдоха, выдоха и пневмотаксиса, расположенными в продолговатом мозге и варолиевом мосту. Нарушение этого отдела вызывает остановку дыхания. Регуляторный (высший) отдел дыхательного центра расположен в лобной доле коры больших полушарий и позволяет произвольно регулировать легочную вентиляцию (глубину и частоту дыхания). Однако эта произвольная регуляция небезгранична и зависит от функциональной активности рабочего отдела, афферентной импульсации, отражающей состояние внутренней среды (в данном случае рН крови, концентрации СО2 и О2 в крови).
Свойства нервных центров. Физиологические свойства нервных центров определяют характер ответных реакций. Эти свойства в значительной мере связаны с особенностями проведения нервных импульсов через синапсы, соединяющие различные нервные клетки.
1. Одностороннее проведение возбуждения. Обусловлено особенностями проведения возбуждения по синапсу. Медиаторы выделяются в пресинаптическом окончании; рецепторы, взаимодействующие с медиаторами, находятся в постсинаптической мембране.
2. Замедление проведения возбуждения – наличие латентного периода (периода от начала действия сигнала до проявления рефлекторного акта). Обусловлено синаптической задержкой в центральной части рефлекторной дуги, где возбуждение проходит через синапсы. На выделение и диффузию медиатора в синапсе требуется промежуток времени в 1,5-2 мс (синаптическая задержка). Чем больше нейронов в рефлекторной дуге, тем продолжительнее время рефлекса.
3. Рефлекторный ответ зависти от силы и продолжительности действия раздражителя.
4. Суммация возбуждений. В работе нервных центров значительное место занимают процессы пространственной и временнóй суммации подпороговых раздражений. Временнáя суммация наблюдается, если множество слабых импульсов приходит к нейрону по одному и тому же пути через один синапс с коротким интервалом времени. В результате из пресинаптической мембраны выбрасывается множество квантов медиатора, происходит суммирование изменений на постсинаптической мембране, и она деполяризуется до уровня, достаточного для генерации возбуждения. Пространственная суммация связана с суммированием подпороговых потенциалов, возникающих одновременно в разных синапсах одного нейрона. Оба вида суммации происходят в области аксонного холмика, где и генерируется ПД.
5. Низкая лабильность обусловлена сравнительно низкой скоростью развития синаптической передачи нервного импульса в химическом синапсе (около 50-100 импульсов в секунду, что в 5 – 6 раз ниже лабильности аксона).
6. Утомление нервных центров. Длительное повторное раздражение рецептивного поля рефлекса приводит к ослаблению рефлекторной реакции вплоть до полного исчезновения, что называется утомлением. Этот процесс связан с деятельностью синапсов – в последних наступает истощение запасов медиатора, уменьшаются энергетические ресурсы, происходит адаптация постсинаптического рецептора к медиатору.
Разные нервные центры имеют различную скорость утомления. Менее утомляемы центры вегетативной нервной системы, координирующие работу внутренних органов. Значительно более утомляемы центры соматической нервной системы, управляющие произвольной скелетной мускулатурой.
7. Трансформация ритма. Сигналы, которые уходят от нервного центра по своим параметрам (частота, сила) отличаются от таковых пришедших к нервному центру. Нервные клетки обладают свойством изменять частоту передающихся импульсов, т. е. свойством трансформации ритма.
Характер ответного разряда нейрона зависит от свойств раздражителя, а также и от функционального состояния самого нейрона (его мембранного заряда, возбудимости, лабильности). В обычных условиях, чем сильнее раздражение, тем чаще проявляется ответная импульсация.
При высокой возбудимости нейрона (например, после приема кофеина) может возникать учащение импульсации (мультипликация ритма), а при низкой возбудимости (например, при утомлении) происходит урежение ритма, так как несколько приходящих импульсов должны суммироваться, чтобы, наконец, достичь порога возникновения потенциала действия. Эти изменения частоты импульсации могут усиливать или ослаблять ответные реакции организма на внешние раздражения.
8. Нервные центры очень чувствительны к недостатку кислорода. В нервных клетках осуществляется интенсивный обмен веществ, для чего необходима энергия и постоянный приток нужного количества кислорода. Особенно чувствительны к недостатку кислорода нервные клетки коры больших полушарий головного мозга: после 5 – 6 минут кислородного голодания они погибают. У человека даже кратковременное ограничение мозгового кровообращения приводит к потере сознания. Недостаточное снабжение кислородом легче переносят нервные клетки мозгового ствола: их функция восстанавливается через 15 – 20 минут после полного прекращения кровоснабжения. А функция клеток спинного мозга восстанавливается даже после 30 минут отсутствия кровоснабжения.
9. Нервные центры чувствительны к химическим веществам, которые проходят через гематоэнцефалический барьер. Нервные центры обладают специфической реакцией на различные фармакологические вещества, что говорит о своеобразии протекающих в них процессов. Стрихнин возбуждает нервные центры, блокируя работу тормозных синапсов. Хлороформ и эфир сначала возбуждают, а затем подавляют работу нервных центров. Апоморфин возбуждает рвотный центр, цититон и лобелин – дыхательный центр, а морфин угнетает его работу. Коразол возбуждает клетки двигательной зоны коры, вызывая эпилептические судороги.
10. Нервные центры всегда находятся в тонусе. Это обусловлено тем, что они постоянно получают импульсацию от других структур мозга. Кроме того, к нервным центрам постоянно идут импульсы обратной афферентации от рецепторов исполнительных органов. В ответ центры посылают редкие импульсы к органам, поддерживая в них соответствующий тонус. Даже во время сна мышцы не расслабляются полностью и контролируются соответствующими центрами.
11. Последействие нервных центров. Следовые процессы или последействие означает, что после окончания действия раздражителя активное состояние нервного центра продолжается еще некоторое время. Длительность следовых процессов различна. В спинном мозге – несколько секунд или минут. В подкорковых центрах мозга – десятки минут, часы и даже дни. В коре больших полушарий – до нескольких десятков лет.
Следовые процессы имеют важное значение в понимании механизмов памяти. Непродолжительное последействие до 1 часа связано с циркуляцией (реверберацией) импульсов в нервных цепях (Р. Лоренте де Но, 1934) и обеспечивает кратковременную память. Механизмы долговременной памяти основаны на изменении структуры белков. В процессе запоминания, согласно биохимической теории памяти (Х. Хиден, 1969) происходят структурные изменения в молекулах РНК, на основе которых строятся измененные белки с отпечатками прежних раздражителей. Эти белки длительно содержатся в нейронах, а также в глиальных клетках головного мозга.
1. 11 Торможение в ЦНС. Торможение в ЦНС– это активный процесс, проявляющийся в подавлении или ослаблении возбуждения. В отличие от возбуждения, торможение по нервным волокнам не распространяется.
Явление торможения в нервных центрах было впервые открыто И. М. Сеченовым в 1862 г. Значительно позже английский физиолог Шеррингтон открыл, что процессы возбуждения и торможения участвуют в любом рефлекторном акте.
Значение торможения:
1) Координационное – процесс торможения обеспечивает упорядоченность или координацию в работе нервных центров, например, чтобы согнуть руку, надо возбудить центр сгибания, посылающий нервные импульсы на бицепс, и затормозить центр разгибания, посылающий нервные импульсы на трицепс.
2) Охранительное значение – при действии сверхсильных раздражителей в нервном центре развивается не возбуждение, а торможение, в результате восстанавливаются запас АТФ и медиатора.
3) Ограничение притока афферентных импульсов в ЦНС (так как мозг не в состоянии переработать весь объем второстепенной малозначимой для человека информации).
Важнейшая роль торможения в координационной деятельности ЦНС наглядно прослеживается при введении в организм подопытного животного определённой дозы стрихнина, временно лишающего ЦНС способности развивать торможение. У такого животного практически исчезает всякая координация рефлексов: любое локальное раздражение вызывает одновременную двигательную реакцию всей мускулатуры тела.
По месту возникновения различают пресинаптическое торможение и постсинаптическое. При пресинаптическом торможении тормозной эффект реализуется на пресинаптической мембране, этот вид торможения участвует в ограничении притока чувствительных импульсов в мозг. Возникает на аксо-аксональных синапсах.
Постсинаптическое торможение осуществляется на постсинаптической мембране. Это основной вид торможения. Развивается в истинных тормозных синапсах (аксосоматических и аксодендрических) с участием тормозных медиаторов, которые изменяют свойства постсинаптической мембраны таким образом, что способность нервной клетки генерировать процессы возбуждения подавляется.
По нейронной организации торможение подразделяют на поступательное, возвратное, латеральное (боковое) и реципрокное.
1. Поступательное торможение обусловлено включением тормозных нейронов на пути следования возбуждения.
2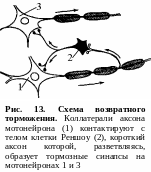 .
Возвратное
торможение осуществляется
вставочными тормозными нейронами
(клетками Реншоу). Импульсы от мотонейронов,
через отходящие от его аксона коллатерали,
активируют клетку Реншоу, которая в
свою очередь вызывает торможение
разрядов данного мотонейрона (рис. 13).
Это торможение реализуется за счет
тормозных синапсов, образованных клеткой
Реншоу на теле активирующего ее
мотонейрона. Таким образом, из двух
нейронов формируется контур с отрицательной
обратной связью, которая дает возможность
стабилизировать частоту разряда
мотонейрона и подавлять избыточную его
активность.
.
Возвратное
торможение осуществляется
вставочными тормозными нейронами
(клетками Реншоу). Импульсы от мотонейронов,
через отходящие от его аксона коллатерали,
активируют клетку Реншоу, которая в
свою очередь вызывает торможение
разрядов данного мотонейрона (рис. 13).
Это торможение реализуется за счет
тормозных синапсов, образованных клеткой
Реншоу на теле активирующего ее
мотонейрона. Таким образом, из двух
нейронов формируется контур с отрицательной
обратной связью, которая дает возможность
стабилизировать частоту разряда
мотонейрона и подавлять избыточную его
активность.
3. Латеральное торможение, т. е. возникает процесс торможения группы нейронов, расположенных рядом с группой возбужденных клеток. Этот вид торможения распространен в сенсорных системах.
4. Реципрокное торможение, т. е. процесс, основанный на том, что сигналы по одним и тем же афферентным путям обеспечивают возбуждение одной группы нейронов, а через вставочные тормозные клетки вызывают торможение другой группы нейронов. Этот тип торможения называют также сопряженным. Проявляется, например, на уровне мотонейронов спинного мозга, иннервирующих мышцы-антагонисты (сгибатели – разгибатели конечностей). При сгибании руки или ноги центры мышц-разгибателей затормаживаются. Рефлекторный акт возможен только при сопряженном торможении мышц-антагонистов. При ходьбе сгибание ноги сопровождается расслаблением разгибателей и, наоборот, при разгибании тормозятся мышцы-сгибатели. Если бы этого не происходило, то возникла бы механическая борьба мышц, судороги, а не приспособительные двигательные акты.
В процессе онтогенеза за счет развития тормозных нейронов формируются тормозные механизмы ЦНС. Ранней их формой является постсинаптическое торможение, позднее формируется пресинаптическое торможение. Благодаря формированию тормозных механизмов существенно ограничивается иррадиация возбуждения в ЦНС, свойственная новорожденным, появляются индукционные отношения. Безусловные рефлексы становятся более точными и локализованными.
1. 12 Координация рефлекторной деятельности. Координация рефлексов – это согласованное взаимодействие нервных центров для обеспечения какого-либо процесса. Каждый рефлекс представляет собой реакцию всей ЦНС и зависит от всей совокупности межцентральных взаимоотношений. Координация функций обеспечивает рефлекторные акты, соответствующие воздействиям внешней среды. Эти рефлекторные акты могут проявляться со стороны различных систем (мышечной, эндокринной, сердечно-сосудистой). К примеру: при беге рефлекторно работают мышцы сгибателей и разгибателей. Повышается артериальное давление, увеличивается просвет сосудов, учащается биение сердца и дыхания. Все это скоординировано. Координация функций определяется характеристикой взаимоотношений между рефлекторными проявлениями со стороны различных систем организма для реализации определённого физиологического акта. Координационные механизмы достигают своего совершенства к 18 – 20 годам.
Основой координации служат следующие особенности ЦНС.
1. Конвергенция – концентрация импульсов на 1 вставочном нейроне. В ЦНС большинство нейронов получают информацию от разных источников благодаря конвергенции. Это обеспечивает пространственную суммацию импульсов и усиление конечного эффекта. Конвергенция на клетках нервного центра разных афферентных входов предопределяет важные интегративные, перерабатывающие информацию функции центральных нейронов, т. е. высокий уровень интеграционных функций.
2. Дивергенция. Способность нейрона устанавливать многочисленные синаптические связи с различными нервными клетками в пределах одного или разных нервных центров называется дивергенцией. Например, центральные окончания аксонов первичного афферентного нейрона образуют синапсы на многих вставочных нейронах. Благодаря этому одна и та же нервная клетка может участвовать в различных нервных реакциях и контролировать большое число других нейронов, что приводит к иррадиации возбуждения.
3. Наличие в ЦНС двух типов вставочных нейронов: возбуждающих и тормозных. Постоянное взаимодействие процессов возбуждения и торможения, как между разными центрами, так и в пределах каждого из них является важнейшей характеристикой их деятельности.
Механизмы координации
1. Иррадиация возбуждения. Механизм иррадиации базируется на дивергенции терминалей афферентных и вставочных нейронов. Это самый древний механизм координации.
Нейроны разных центров связаны между собой многочисленными вставочными нейронами, поэтому при раздражении рецепторов возбуждение может распространяться не только на нейроны центра данного рефлекса, но и на другие нейроны. Распространение процессов возбуждения на другие нервные центры называют явлением иррадиации. Чем сильнее и длительнее афферентное раздражение и чем выше возбудимость окружающих нейронов, тем больше нейронов охватывает процесс иррадиации. Процессы торможения ограничивают иррадиацию и способствуют концентрации возбуждения в исходном пункте ЦНС.
Процесс иррадиации играет важную положительную роль при формировании новых реакций организма (ориентировочных реакций, условных рефлексов). Благодаря иррадиации возбуждения между различными нервными центрами возникают новые функциональные связи – условные рефлексы. Иррадиация возбуждения может оказать отрицательное воздействие на состояние и поведение организма, нарушая тонкие взаимоотношения между возбужденными и заторможенными нервными центрами и вызывая нарушения координации движений.
2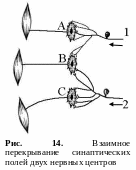 .
Облегчение
и окклюзия.
Конвергенция множества путей, т. е.
нервных цепочек на одной группе
мотонейронов лежит в основе феноменов
облегчения и окклюзии. Облегчение и
окклюзия возможны в том случае, когда
на какой-то части нейронов сходятся
(конвергируют) пресинаптические волокна
из различных афферентных источников
(рис. 14).
.
Облегчение
и окклюзия.
Конвергенция множества путей, т. е.
нервных цепочек на одной группе
мотонейронов лежит в основе феноменов
облегчения и окклюзии. Облегчение и
окклюзия возможны в том случае, когда
на какой-то части нейронов сходятся
(конвергируют) пресинаптические волокна
из различных афферентных источников
(рис. 14).
Облегчение – это превышение эффекта одновременного действия двух слабых пороговых раздражителей над суммой их раздельных эффектов. Например, нейроны А и С получают от каждого афферентного нейрона (1 и 2) достаточное количество синаптических окончаний (например, по 4), чтобы в них возникло возбуждение (рис. 14). Нейрон В получает от тех же афферентных нейронов (1 и 2) меньшее число окончаний (по 2), поэтому их импульсов будет недостаточно, чтобы генерировать в этом нейроне нервный импульс, а возникает лишь подпороговое возбуждение. Вследствие этого, при раздельном раздражении афферентных нейронов 1 и 2 возникают рефлекторные реакции только в нейронах А и С. Но при одновременном раздражении афферентных нейронов 1 и 2 нервный импульс генерируется и нейроном В (происходит суммация нервных импульсов от двух источников). Поэтому выраженность такого суммарного рефлекторного ответа будет больше. Это явление получило название центрального облегчения.
Окклюзия (закупорка) – это явление, противоположное облегчению. Окклюзия возникает при действии сильных надпороговых раздражителей. При взаимном перекрывании синаптических полей двух нервных центров суммарный результат оказывается меньшим, чем сумма отдельных реакций. Так, при раздражении афферентного поля первого нервного центра возникает возбуждение в двух нейронах (А и В), при активации второго центра также возникает возбуждение в двух центрах (С и В), однако при одновременной стимуляции обоих центров (1 и 2) суммарный двигательный ответ равен только трем единицам (возбуждаются центры А, В и С), а не четырем (рис. 14). Это обусловлено тем, что один и тот же мотонейрон (В) относится одновременно к обоим нервным центрам. Явление окклюзии приводит к снижению силы суммарной ответной реакции.
3. Принцип общего конечного пути. Афферентных нейронов в центральной нервной системе в несколько раз больше, чем эфферентных. В связи с этим многие афферентные влияния поступают к одним и тем же вставочным и эфферентным нейронам, которые являются для них общими конечными путями к рабочим органам. Система реагирующих нейронов образует, таким образом, как бы воронку («воронка Шеррингтона»). Множество разнообразных раздражений может возбудить одни и те же мотонейроны спинного мозга и вызвать одну и ту же двигательную реакцию. Так, например, мотонейроны, иннервирующие дыхательную мускулатуру, помимо обеспечения вдоха участвуют в таких рефлекторных реакциях, как чихание, кашель и др.
Английский физиолог Ч. Шеррингтон, установивший принцип общего конечного пути, предложил различать союзные и антагонистические рефлексы. Встречаясь на общих конечных путях, союзные рефлексы взаимно усиливают друг друга, а антагонистические – тормозят. В первом случае в нейронах общего конечного пути имеет место пространственная суммация (например, сгибательный рефлекс усиливается при одновременном раздражении нескольких участков кожи). Во втором случае происходит борьба конкурирующих влияний за обладание общим конечным путем, в результате чего один рефлекс осуществляется, а другие затормаживаются. При этом освоенные движения выполняются с меньшим трудом, так как в их основе лежат упорядоченные во времени синхронизированные потоки импульсов, которые проходят через конечные пути легче, чем импульсы, поступающие в случайном порядке.
Преобладание на конечных путях той или иной рефлекторной реакции обусловлено ее значением для жизнедеятельности организма в данный момент.
В таком отборе важную роль играет наличие в центральной нервной системе доминанты. Она обеспечивает протекание главной реакции. Например, ритмический шагательный рефлекс и одиночный, непрерывный рефлекс сгибания при болевом раздражении являются антагонистическими. Однако спортсмен, внезапно получивший травму, может продолжать бег к финишу, т. е. осуществлять ритмический рефлекс и подавлять болевые раздражения, которые, поступая к мотонейронам сгибательных мышц, препятствуют попеременному сгибанию и разгибанию ноги.
4. Обратная связь или вторичная афферентация. Всякий двигательный акт, вызванный афферентным раздражителем, сопровождается возбуждением рецепторов мышц, сухожилий, суставных сумок. Сигналы с проприорецепторов вторично поступают в ЦНС, что позволяет осуществлять коррекцию ее деятельности и саморегуляцию в соответствии с текущими потребностями организма и наличной обстановкой. Этот важный принцип рефлекторной саморегуляции функций организма называется принципом обратной связи.
Кроме того, за счет обратной связи поддерживается тонус нервных центров.
5. Реципрокные (сопряженные) взаимоотношения между нервными центрами. В основе взаимосвязи между нервными центрами лежит процесс индукции – развитие (индуцирование) противоположного процесса. Индукция ограничивает распространение (иррадиацию) нервных процессов и обеспечивает концентрацию.
Различают одновременную и последовательную индукцию. Сильный процесс возбуждения в нервном центре вызывает (индуцирует) торможение в соседних нервных центрах (одновременная отрицательная индукция), а сильный тормозной процесс индуцирует в соседних нервных центрах возбуждение (одновременная положительная индукция). Так, для сгибательного движения в суставе необходимо не только сокращение мышц-сгибателей, но и одновременное расслабление мышц- разгибателей. При этом в мотонейронах мышц-сгибателей возникает процесс возбуждения, а в мотонейронах мышц-разгибателей – процесс торможения. При возбуждении же центров разгибателей, наоборот, тормозятся центры сгибателей.
При смене процессов возбуждения и торможения в пределах одного центра говорят о последовательной отрицательной или положительной индукции. Она имеет большое значение при организации ритмической двигательной деятельности, обеспечивая попеременное сокращение и расслабление мышц.
Установлено, что у человека во время ходьбы и бега основной формой координации являются реципрокные отношения, но помимо них имеются фазы одновременной активности мышц-антагонистов голеностопного и особенно коленного и тазобедренного суставов. Длительность фаз одновременной активности увеличивается с повышением скорости перемещения.
У детей четкие индукционные взаимоотношения между процессами торможения и возбуждения начинают развиваться в возрасте с 3-х до 5-и лет, так как в этом возрасте возрастает сила нервных процессов.
6. Доминанта. Активность нервных центров непостоянна, и преобладание активности одних из них над активностью других вызывает заметные перестройки в процессах координации рефлекторных реакций. В 1923 г. А. А. Ухтомский сформулировал принцип доминанты как рабочий принцип деятельности нервных центров.
Доминанта – это временное преобладание одного нервного центра или группы центров над другими, определяющее текущую деятельность организма.
Основные черты, доминанты следующие: 1) повышенная возбудимость нервных центров; 2) стойкость возбуждения во времени; 3) способность усиливать свое возбуждение за счет суммации нервных импульсов, идущих в другие центры (в результате этого раздражение различных рецепторных полей начинает вызывать рефлекторный ответ, характерный для деятельности этого доминантного центра); 4) доминирующий центр по механизму одновременной индукции вытормаживает другие центры.
Доминантный очаг в ЦНС может возникать под влиянием разных факторов, в частности, сильной афферентной стимуляции, гормональных воздействий, изменения химизма крови, мотиваций и т. д.
Несмотря на относительную стойкость и инертность возбуждения в доминантном очаге, деятельность ЦНС в нормальных условиях существования весьма динамична и изменчива. ЦНС обладает способностью к перестройке доминантных отношений в соответствии с изменяющимися потребностями организма, и в течение всей жизни человека одна доминанта сменяет другую. Сильные доминанты остаются даже во сне.
Доминантный очаг у ребенка возникает быстрее и легче, чем у взрослых, но для него характерна низкая устойчивость к внешним раздражителям. С этим в значительной степени связана неустойчивость внимания у детей: новые раздражители легко вызывают новую доминанту.
7. Пластичность нервных центров. Под пластичностью понимают функциональную изменчивость и приспособляемость нервных центров, их способность выполнять новые, необычные для них рефлекторные акты. Это особо ярко проявляется после удаления различных отделов мозга. Если были частично удалены какие-то отделы мозжечка или коры больших полушарий, нарушенная функция со временем может восстановиться.