

20. Окислительное дезаминирование глутаминовой кислоты.
Глутаматдегидрогеназа.
ЭТО ЕСТЬ обратимая реакция и при повышении концентрации аммиака в клетке может протекать в обратном направлении, как восстановительное аминирование α-кетоглутарата.
Глутаматдегидрогеназа очень активна в митохондриях клеток практически всех органов, кроме мышц. Этот фермент - олигомер, состоящий из 6 субъединиц. Глутаматдегидрогеназа играет важную роль, так как является регуляторным ферментом аминокислотного обмена. Аллостерические ингибиторы глутаматдегидрогеназы (АТФ, ГТФ, NADH) вызывают диссоциацию фермента и потерю глутаматдегидрогеназной активности. Высокие концентрации АДФ активируют фермент. Таким образом, низкий энергетический уровень в клетках стимулирует разрушение аминокислот и образование α-кетоглутарата, поступающего в ЦТК как энергетический субстрат. Синтез глутаматдегидрогеназы может индуцироваться стероидными гормонами (кортизолом).
21. Непрямое дезаминирование, транс-дезаминирование, химизм, био-
логическая роль.
Большинство аминокислот не способно де-заминироваться в одну стадию, подобно Глу. Аминогруппы таких аминокислот в результате трансаминирования переносятся на α-кетоглу-тарат с образованием глутаминовой кислоты, которая затем подвергается прямому окислительному дезаминированию. Такой механизм дезаминирования аминокислот в 2 стадии получил название трансдезаминирования, или непрямого дезаминирования.

Непрямое дезаминирование аминокислот происходит при участии 2 ферментов: амино-трансферазы (кофермент ПФ) и глутаматдегид-рогеназы (кофермент NAD+).
Значение этих реакций в обмене аминокислот очень велико, так как непрямое деза-минирование - основной способ дезаминирования большинства аминокислот. Обе стадии непрямого дезаминирования обратимы , что обеспечивает как катаболизм аминокислот, так и возможность образования практически любой аминокислоты из соответствующей α-кетокислоты.
22. Декарбоксилирование аминокислот.
Декарбоксилирование - процесс отщепления карбоксильной группы аминокислот в виде СО2. Зачем нам это надо? В результате этих процессов в животных тканях образуются биогенные амины.
Реакции декарбоксилирования необратимы катализируются ферментами декарбоксилаза. Их простетическая группа - пиридоксальфосфат.
Механизм реакции напоминают РЕАКЦИЮ трансаминирования с участием пиридоксальфостфат и также осущестляется путем образования шиффова основания ПФ и аминокислоты.
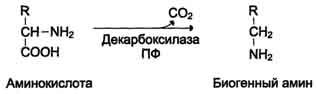
23. Биогенные амины, происхождение, функции.
Что такое биогенные амины? Это амины, образовавшиевся при декарбоксилировании аминокислот! Они играют ряд физиологически ролей:
- нейромедиаторы (серотонин, дофамин, ГАМК и др.);
- гормоны (норадреналин, адреналин);
- регуляторные факторы местного действия (гистамин, карнозин, спермин и др.);
Накопление биогенных амино может сказаться паршивым образом на физиологическом статусе и вызывать нарушения функций в организме. Затим, есть ряд механизмов по их обезвреживанию, и усё сводится к окислительному дезаминированию этих аминов с образование соответствующих альдегидов и освобождению аммиака.
Ферменты, катализирующие эту реакцию называются моноамин- и диаминоксидазы.
И сноаа, две стадии:
- анаэробная стадия, характеризуется образованием альдегида, аммиака и восстановленного фермента.
- Затем фермент окисляет атомарным кислородом, образуется перекись водорода, которая распадает на воду и ксилород.