

46. Распад нуклеиновых кислот, нуклеазы пищеварительного тракта и
тканей.
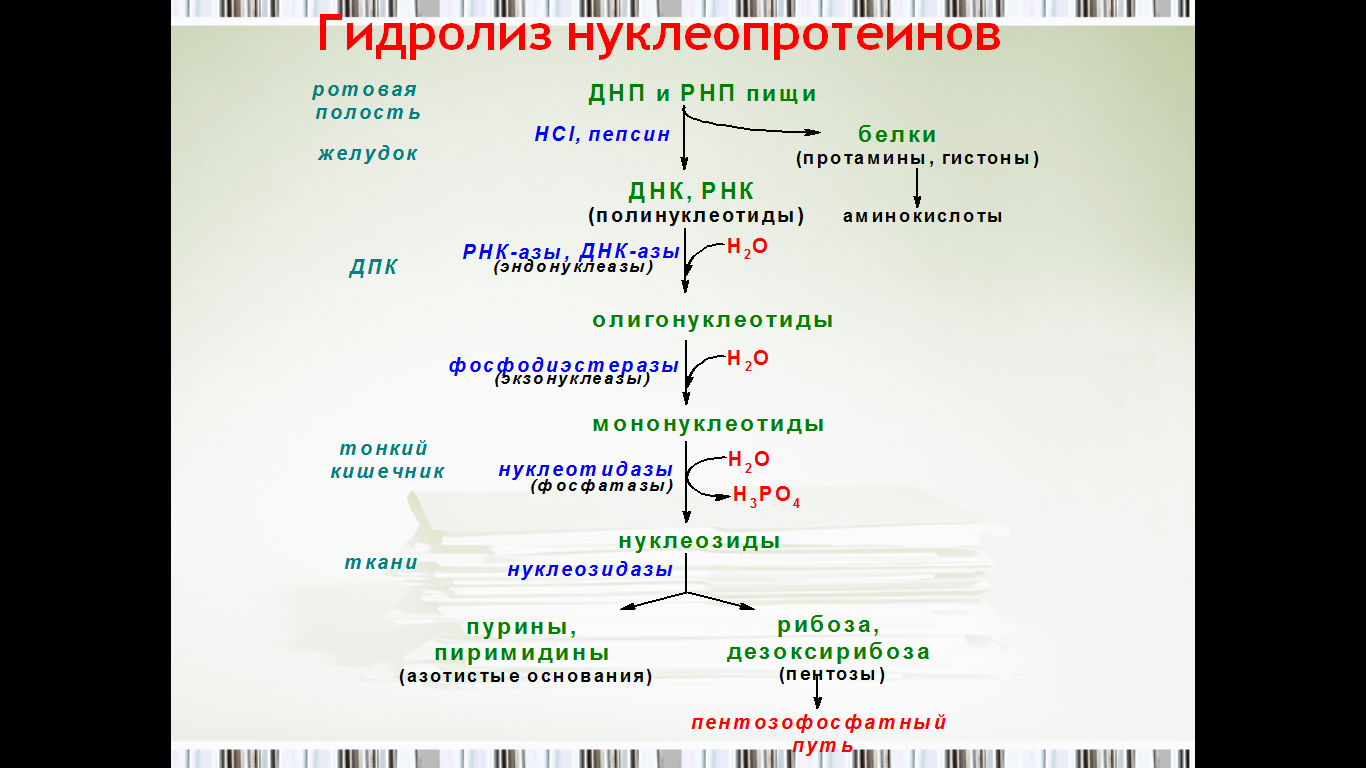
Схема говорит о многом. Съели курочку, её надо разобрать. ДНП и РНП чрез пищевод попадает в желудок, там, под действием агрессивных агентов - соляной кислоты и пепсинов расщепляются до полинуклеотидов. Дальше в ход вступают эндонуклеазы, образуются олигонуклеотиды и экзонуклеазы, ура, получили мононуклеотиды.
Продукт мигрирует тонкий кишечник, нуклеотидазы синтезируют нуклеозиды.
Парам-пам-пам, в тканях нуклеозидазы образуют строй материалы - азотистые основания и пентозы (а их могут пустить пентозофосфатный путь) Усё.
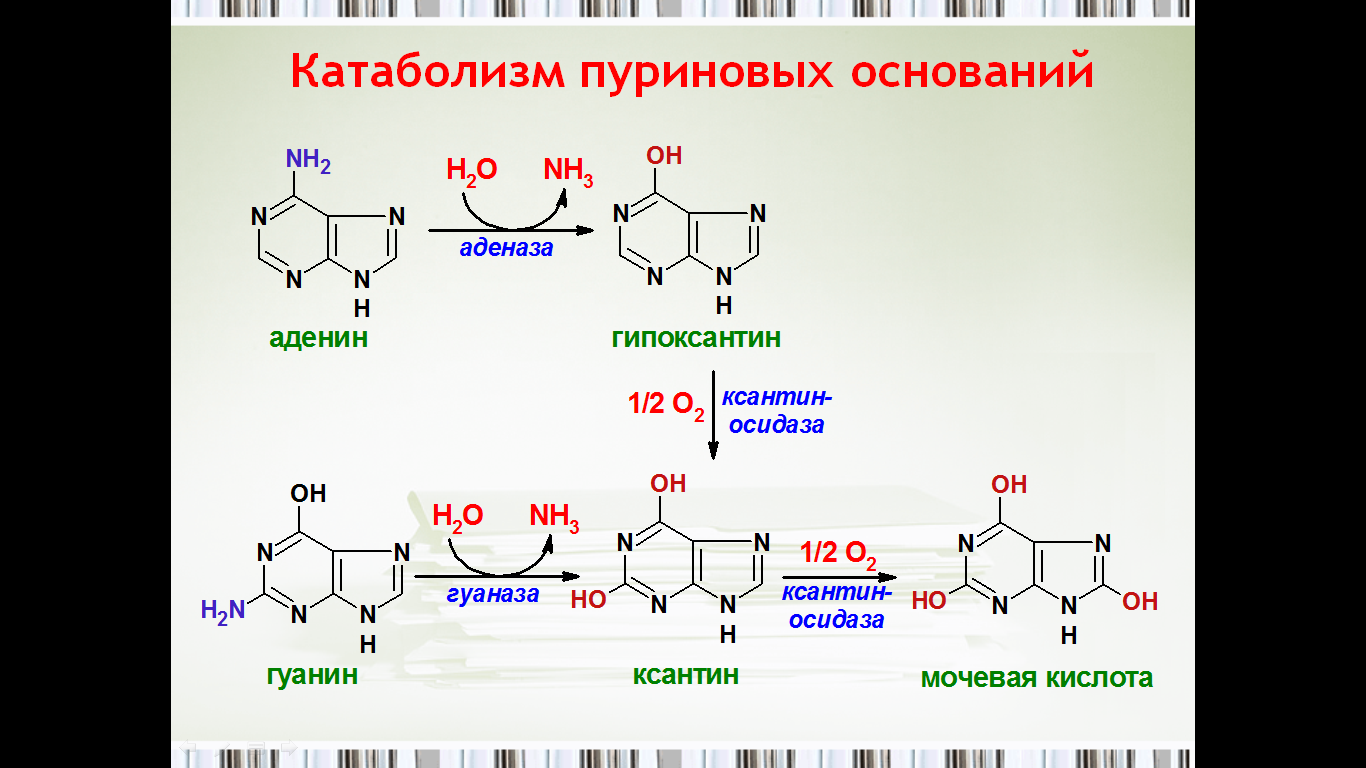
47. Распад пуриновых нуклеотидов.
48. Биосинтез пуриновых нуклеотидов, происхождение атомов «с» и «n» в пуриновом кольце.


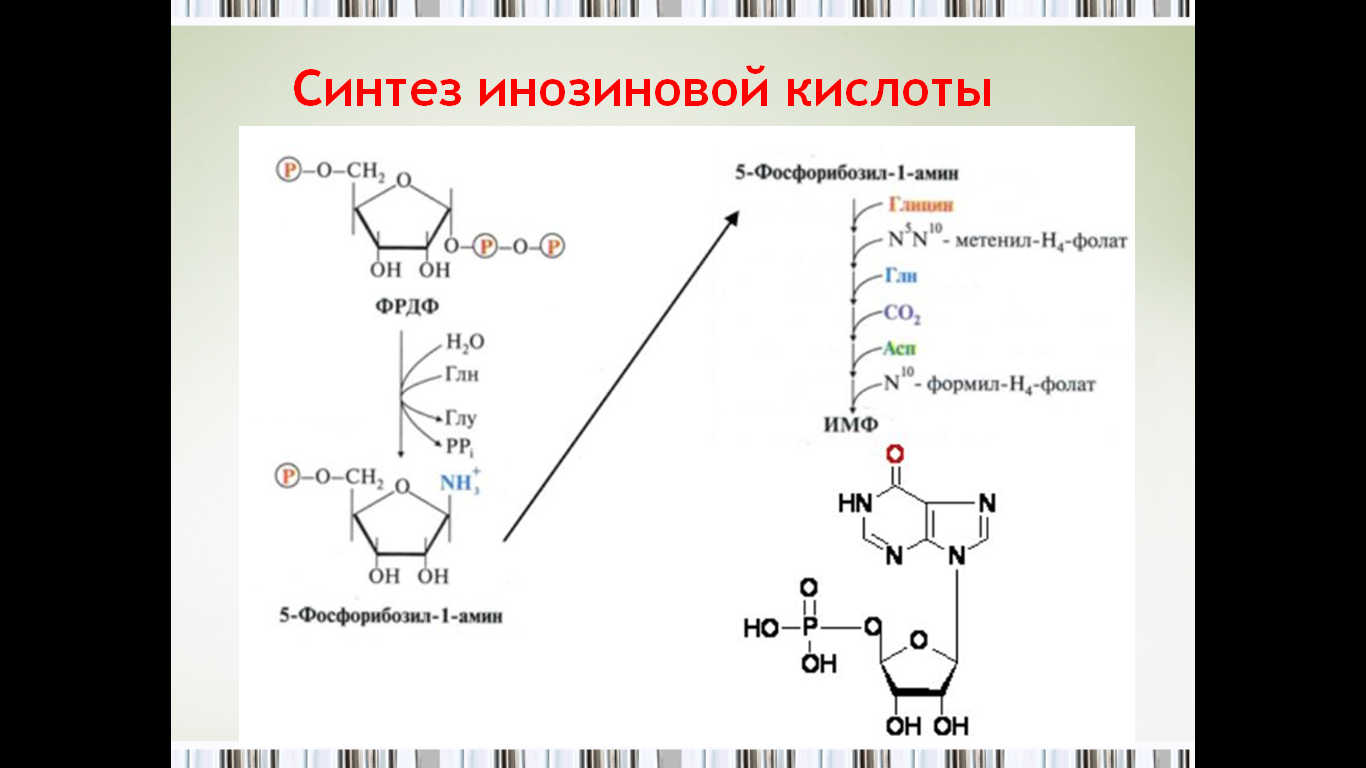
49. Инозиновая кислота как предшественник пуриновых мононуклеотидов.
Образование пуриновых нуклеотидов (адениловой и гуаниловой кислот, АМФ и ГМФ) осуществляется из инозиновой кислоты (ИМФ). Причем в синтезе обоих мононуклеотидов участвуют по два фермента, отличных по своему механизму действия. Образование ГМФ из ИМФ катализируют ИМФ-дегидрогеназа и ГМФ-синтетаза, а образование АМФ из того же предшественника катализируется последовательным действием аденилосукцинатсинтетазы и аденилосукцинатлиазы.
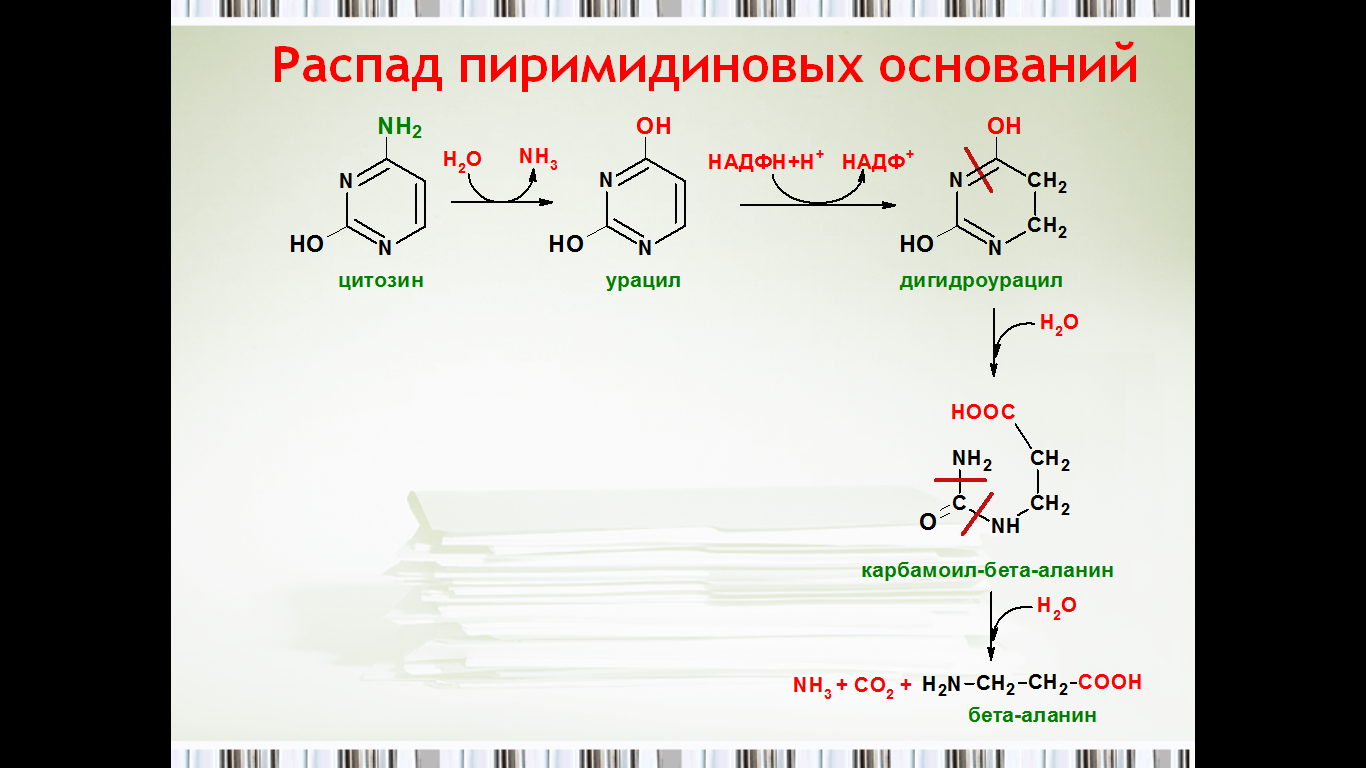
50. Распад пиримидиновых нуклеотидов.
51. Биосинтез пиримидиновых нуклеотидов.

52. Регуляция биосинтеза пуриновых и пиримидиновых мононуклеотидов.
Регуляция синтеза пиримидиновых нуклеотидов
Регуляторным ферментом в синтезе пири-мидиновых нуклеотидов является полифункциональный КАД-фермент. УМФ и УТФ ал-лостерически ингибируют, а ФРДФ активирует его карбамоилсинтетазную активность, тогда как активность аспартаттранскарбамоилазного домена ингибирует ЦТФ, но активирует АТФ
Этот способ регуляции позволяет предотвратить избыточный синтез не только УМФ, но и всех других пиримидиновых нуклеотидов и обеспечить сбалансированное образование всех четырёх основных пуриновых и пиримидиновых нуклеотидов, необходимых для синтеза РНК.
Регуляция синтеза пуиновых нуклеотидов
Основным показателем, от которого зависит синтез пуриновых нуклеотидов, служит концентрация ФРДФ, которая, в свою очередь, зависит от скорости его синтеза, утилизации и разрушения. Количество ФРДФ определяется доступностью рибозо-5-фосфата и активностью ФРДФ синтетазы - фермента, чувствительного к концентрации фосфата и пуриновых нуклеотидов.
Внутриклеточная концентрация ФРДФ строго регулируется и обычно низкая. ФРДФ синтета-за - аллостерический фермент. Он активируется неорганическим фосфатом (Pi) и ингибируется пуриновыми нуклеозидмоно-, ди- и трифосфатами, которые по эффективности ингибирования распределяются в следующем порядке: НМФ > НДФ > НТФ. ФРДФ служит не только субстратом, но и аллостерическим активатором второй реакции синтеза пури-нонуклеотидов de novo, которую катализирует амидофосфорибозилтрансфераза.
Пуриновые нуклеотиды, особенно АМФ и ГМФ по механизму отрицательной обратной связи ингибируют амидофосфорибозилтрансфе-разу, которая катализирует первую специфическую реакцию синтеза пуриновых нуклеотидов de novo.