

Пример расчета демографических показателей в гипотетической стабильной популяции с дискретными возрастными классами (по Пианке, 1981)
Возраст (x) |
lx |
mx |
lx mx |
x lx mx |
Ex (см.ниже) |
Vx (см.ниже) |
0 |
1,0 |
0,0 |
0,0 |
0,00 |
3,40 |
1,00 |
1 |
0,8 |
0,2 |
0,16 |
0,16 |
3,00 |
1,25 |
2 |
0,6 |
0,3 |
0,18 |
0,36 |
2,67 |
1,40 |
3 |
0,4 |
1,0 |
0,40 |
1,20 |
2,50 |
1,65 |
4 |
0,4 |
0,6 |
0,24 |
0,96 |
1,50 |
0,65 |
5 |
0,2 |
0,1 |
0,02 |
0,10 |
1,00 |
0,10 |
6 |
0,0 |
0,0 |
0,00 |
0,00 |
0,00 |
0,00 |
Суммы |
|
2,2 (валовая рождаемость) |
1,00 (R0) |
2,78 (T) |
|
|
Ожидаемая продолжительность жизни:
Е0=(l0+l1+l2+l3+l4+l5)/ l0=(1,0+0,8+0,6+0,4+0,4+0,2)/1,0=3,4/1,0;
Е1=(l1+l2+l3+l4+l5)/ l1=(0,8+0,6+0,4+0,4+0,2)/0,8=2,4/0,8=3,0;
Е2=(l2+l3+l4+l5)/ l2=(0,6+0,4+0,4+0,4+0,2)/0,6=1,6/0,6=2,67;
Е3=(l3+l4+l5)/ l3=(0,4+0,4+0,2)/0,4=1,0/0,4=2,5;
Е4=(l4+l5)/ l4=(0,4+0,2)/0,4=0,6/0,4=1,5;
Е5=l5/l5=0,2/0,2=1,0.
Репродуктивная ценность:
V0=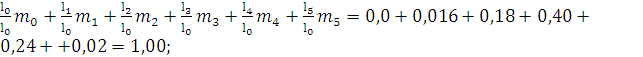
V1=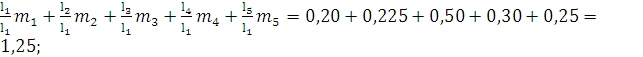
V2=![]()
V3=![]()
V4=![]()
V4=![]()
Начальный, или «нулевой», возраст выбирается достаточно условно. Например, при изучении птиц за начальный возраст принимают либо момент откладки яиц, либо момент вылупления птенцов, либо момент вылета птенцов из гнезда. У планктонных рачков и коловраток за начальный возраст принимают либо момент появления яйца, либо момент выхода молоди из яйца. Выделение возрастных классов производится в зависимости от продолжительности жизни. Для человека, например, выбирается интервал в 5 лет, дли мелких грызунов – месяц или несколько месяцев, для многих насекомых – около недели, для планктонных ракообразных (ветвистоусые рачки) – от одного до нескольких дней и т.д. В том случае, если продолжительность жизни животного или растения очень велика (превышает продолжительность жизни человека) или нет возможности определить начальный возраст, исследователь анализирует возрастную структуру отдельных возрастных групп и, зная численность этих групп, рассчитывает специфичную для каждого возраста смертность. Полученный материал представляется в статистической таблице повозрастной смертности, в отличие от когортных таблиц выживания или таблиц жизни.
Доля особей, доживших до возраста х, обозначается 1х и соответствует средней вероятности доживания новорожденной особи до данного возраста. На основании полученных таким образом значений числа доживших до определенного возраста особей строятся кривые выживаемости. По оси ординат обычно откладывают логарифм числа доживших особей, по оси абсцисс – возраст. Нередко существует специфика выживаемости, определяемая полом животных, что свойственно и человеческим популяциям. В таких случаях строят кривые выживаемости для самцов и самок отдельно.
Первая попытка составления таблицы выживаемости принадлежит основателю демографии Д. Гранту (1620–1674). Исходным материалом для его расчетов служили данные о смертности жителей Лондона, собранные в церковных приходах с сугубо практической целью – сообщать о начале эпидемии чумы. Так как в то время динамика населения Лондона определялась в значительной степени иммиграцией из сельской местности, которая не учитывалась, полученные данные оказались трудными для интерпретации. В 1693 г. Э. Галлей, известный по описанной им комете, составил таблицу выживаемости населения для города Бреслау (Вроцлава), в котором численность населения была стационарна.
В экологию таблицы жизни введены Р. Перлом в 1921 г. В 1928 г. Перл описал три основных типа кривых выживаемости (рис. 65).
Первый тип отражает ситуацию, при которой смертность до определенного возраста низкая, а затем с увеличением возраста быстро возрастает. Такой тип характерен для имаго многих насекомых, ящериц Xantusia vigilis и Scincella laterale, барана Далла, африканских копытных, вероятно, для большинства млекопитающих, в том числе и человека.
Второй тип свидетельствует об относительном постоянстве смертности всех возрастов. Этот тип свойствен преимущественно птицам, но встречается и у многих других животных, например, у гидры, некоторых ящериц (Uta stansburiana, Eumeces fasciatus), бородавочника Phacochoerus aethiopicus.
Третий тип, или так называемый «тип устрицы» (Ostrea), характеризует очень высокую смертность в раннем возрасте. Свойствен видам с большой плодовитостью и отсутствием заботы о потомстве, к числу которых относятся практически все морские и пресноводные беспозвоночные, рыбы, а также большинство насекомых, если рассматривать не имаго, а весь онтогенез.
На рисунке 65 представлены еще два типа кривых, близких к типу 2 и нередко встречающихся в популяциях животных, для которых обычен тип 2.
На рис. 65, а показано существенное различие выживаемости самцов и самок буйвола, что характерно и для ряда других млекопитающих.
По данным таблиц жизни легко рассчитать как ожидаемую продолжительность жизни различных возрастных групп популяции, так и репродуктивную ценность (табл. 34, рис. 65).
Ожидаемая продолжительность жизни (Ех) в возрасте х – это средняя продолжительность оставшейся жизни особей, доживших до возраста х. Естественно, что средняя продолжительность жизни будет средней величиной от значений ожидаемой продолжительности жизни всех возрастов.

Рис 65. Основные типы кривых выживаемости:
а – кривые выживаемости самок (1) и самцов (2) в популяции буйвола;
б – выпуклая кривая (1); диагональ (2); вогнутая кривая (3); кривые, близкие к кривой 2 (4,5)
В этой же таблице представлена величина тх – число потомков, производимых в среднем особью возраста х за данный возрастной период, или повозрастная рождаемость. Сумма тх для всех возрастов называется валовой рождаемостью (gross reproductive rate). Таблицы тх сильно варьируются в зависимости от условий среды и биологических особенностей видов, как и таблицы 1х.
На основании данных, приведенных в таблицах жизни, рассчитываются и важнейшие показатели скорости популяционного роста: чистое воспроизведение, или чистая скорость размножения (Ro), время генерации (T) и врожденная скорость популяционного роста (rmax ).
Чистая скорость размножения (R0) (net reproductive rate) – это среднее число потомков, произведенных средней особью когорты за всю жизнь. Расчет производим по формуле:
R0=∑ lx mx
Популяция растет, если Ro> 1, стационарна, если R0= 1, и сокращается, если Rq< 1.
Размножение может происходить один раз за жизнь и многократно в течение жизни. Обычно возраст первого размножения определяется как α, возраст последнего – как ω. Время генерации или продолжительность поколения (T) для организмов, размножающихся только один раз, равно времени от рождения до размножения. Для многократно размножающихся животных T рассчитывается по формуле:
T=∑ lx mx x / ∑ lx mx
Из таблиц жизни рассчитывается и так называемая врожденная (intrinsic), или мгновенная (instantaneous), скорость популяционного роста (rтах), которая является мерой мгновенной удельной (в расчете на особь) скорости изменения размера популяции и выражает число особей, отрождаемых в популяции в единицу времени на одну особь. Если повозрастная рождаемость и выживаемость стабильны и последовательных генерациях, т.е. нет ограничения роста, можно рассчитать количество особей через промежуток времени, равный Т, по формуле:
NT=NoerT.
Известно, что NT /N0= R0. Значит, Ro = erT. Отсюда rmax =lnR0 /T.
Необходимо знать, что величина rmax, во–первых, имеет размерность «время –1»; во–вторых, определяется совокупностью рождаемости и смертности всех возрастов; в–третьих, значение rmax не зависит от плотности популяции.
В табл. 35 приведены значения rтах и T для различных групп организмов от бактерий до млекопитающих. Как видим, увеличение массы организма, как правило, сопровождается уменьшением rтах и увеличением Т.
В замкнутой популяции скорость популяционного роста r определяется как разница между мгновенной рождаемостью (b) и мгновенной смертностью (d). Когда значение Ro максимально высоко, тогда и скорость популяционного роста будет максимальной.
В естественных популяциях постоянство рождаемости и смертности возрастов встречается крайне редко из–за изменчивости биотических и абиотических факторов, влияющих на рост. Методы расчета этих показателей в природных популяциях рассмотрены в главе, посвященной динамике природных популяций.
Благодаря способности к воспроизведению популяция увеличивает численность. Ч. Кребс (Krebs, 1994) рассматривает два аспекта воспроизведения (reproduction): фертильность (fertility) как физиологическую способность давать потомство (здесь более подходит термин «воспроизводительная способность») и скорость воспроизведения (fecundity) как экологический показатель количества новорожденных, производимых в единицу времени. Следует различать потенциальную и реализованную скорость воспроизведения. Так, реализованная скорость воспроизведения для человеческой популяции – одно рождение в 15 лет в детородном возрасте, тогда как потенциальная – одно рождение на 10–11 месяцев в детородном возрасте.
Таблица 35.