

Вопросы по микробиологии.
3)Виды микроскопии
Световая микроскопия
При использовании этого метода исследователь оперирует следующими понятиями:
Увеличение – физическое свойство линз объектива и окуляра. Увеличение микроскопа оценивают как произведение увеличения объектива и увеличения окуляра.
Минимальный размер наблюдаемого объекта (d) и разрешение микроскопа – значения, зависящие от характеристик линз объектива, длины волны и от коэффициента преломления среды, отделяющей изучаемый объект от линз объектива или конденсора. Увеличивают разрешение микроскопа применением жидких сред (иммерсионные среды), т.к. коэффициент их преломления больше коэффициента преломления воздуха. В микроскопии используют масляную, глицериновую и водную иммерсионные среды. Теоретически возможный предел разрешения светового микроскопа – 0,2 мкм (минимальное расстояние, на котором различимы два объекта).
Специальные виды микроскопии
Темнопольная. Используют специальный конденсор, выделяющий контрастирующие структуры неокрашенного материала. Темнопольная микроскопия позволяет наблюдать живые объекты. Наблюдаемый объект выглядит как освещенный на темном поле. При этом лучи от осветителя падают на объект сбоку, а в линзы микроскопа поступают только рассеянные лучи.
Фазово-контрастная микроскопия позволяет изучать живые и неокрашенные объекты. При прохождении света через окрашенные объекты изменяется амплитуда световой волны, а при прохождении света через неокрашенные – фаза световой волны, что и используют для получения высококонтрастного изображения в фазово-контрастной и интерференционной микроскопии.
Поляризационная микроскопия - формирование изображения неокрашенных анизотропных структур (например, коллагеновые волокна и миофибриллы).
Интерференционная микроскопия объединяет принципы фазово-контрастной и поляризационной микроскопии и применяется для получения контрастного изображения неокрашенных объектов.
Люминесцентная микроскопия применяется для наблюдения флюоресцирующих (люминесцирующих) объектов. В люминесцентном микроскопе свет от мощного источника проходит через два фильтра. Один фильтр задерживает свет перед образцом и пропускает свет длины волны, возбуждающей флюоресценцию образца. Другой фильтр пропускает свет длины волны, излучаемой флуоресцирующим объектом. Таким образом, флюоресцирующие объекты поглощают свет одной длины волны и излучают в другой области спектра.
Флюоресцирующие красители (флюоресцин, родамин и др.) избирательно связываются со специфическими макромолекулами.
Электронная микроскопия
Теоретическое разрешение просвечивающего ЭМ составляет 0,002 нм. Реальное разрешение современных микроскопов приближается к 0,1 нм. Для биологических объектов разрешение ЭМ на практике составляет 2 нм.
Просвечивающий ЭМ состоит из колонны, через которую в вакууме проходят электроны, излучаемые катодной нитью. Пучок электронов, фокусируемый кольцевыми магнитами, проходит через подготовленный образец. Характер рассеивания электронов зависит от плотности образца. Проходящие через образец электроны фокусируют, наблюдают на флюоресцирующем экране и регистрируют при помощи фотопластинки.
Сканирующий ЭМ применяют для получения трехмерного изображения поверхности исследуемого объекта.
Метод сколов (замораживания-скалывания) применяют для изучения внутреннего строения клеточных мембран. Клетки замораживают при температуре жидкого азота в присутствии криопротектора и используют для изготовления сколов. Плоскости скола проходят через гидрофобную середину двойного слоя липидов. Обнаженную внутреннюю поверхность мембран оттеняют платиной, полученные реплики изучают в сканирующем электронном микроскопе.
4)Морфология М/О
Некоторые виды бактерий образуют споры и капсулы (рис. 10). Капсула является продуктом набухания и ослизнения оболочки клетки; она предохраняет бактерии от влияния неблагоприятных факторов. При неблагоприятных условиях внутри некоторых палочковидных бактерий образуются округлые тельца - споры.
Спорообразующие палочки (бациллы) могут существовать в двух формах: вегетативной, т.е. способной к росту и размножению, и споровой, неспособной к размножению. Спора представляет собой микробную клетку, потерявшую большое количество воды и покрывшуюся плотной оболочкой. Внутри микробной клетки образуется только одна спора, которая служит для сохранения вида. Если диаметр спор превышает поперечник микробной клетки, - это клостридии (например, возбудитель столбняка).
К микроорганизмам относят микроскопические живые существа, не образующие хлорофилл, в том числе бактерии, грибы (плесени, дрожжи, актиномицеты).
Большинство микробов одноклеточные и лишь некоторые из них являются многоклеточными. К группе одноклеточных относятся бактерии, простейшие, дрожжи, отдельные виды плесневых грибов, а к группе многоклеточных - нитчатые бактерии и большинство плесеней. Вирусы не имеют клеточной структуры в отличие от других микроорганизмов.
Бактерии. Форма и размер бактерий. По внешнему виду различают три основные формы бактерий: шаровидную (кокки), палочковидную (цилиндрические) и извитую (рис. 8).
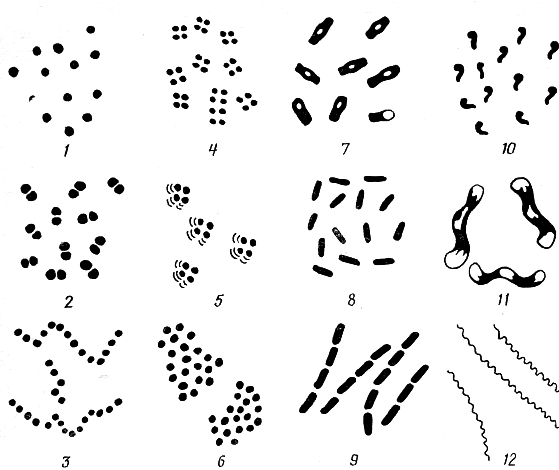 Рис.
8. Основные формы бактерий:
Рис.
8. Основные формы бактерий:
1 — микрококки; 2 — диплококки; 3 — стрептококки; 4 — тетракокки; 5 -сарцины; 6 — стафилококки; 7 — бациллы; 8 — бактерии; 9 — стрептобактерии; 10 — вибрионы; 11 — спириллы; 12 — спирохеты
Размеры бактерий могут колебаться в зависимости от условий обитания и влияния внешней среды (питание, температура, влажность и др.). Размер кокковидных форм колеблется в пределах от 0,75 до 2 мкм, палочковидных от 0,3-1 до 2-10 и извитых от 0,1-0,15 до 3-20 мкм.
Кокки - большинство их имеет правильную форму шара, но некоторые виды вытянуты и напоминают свечу, ланцет, бобы. В зависимости от взаимного расположения клеток (после деления) кокки подразделяются на микрококки - одиночные, беспорядочно расположенные кокки; диплококки - располагаются попарно; стрептококки - образуют цепочку при делении кокков в одной плоскости; тетракокки - сочетания по четыре кокка; сарцины - кокки, соединенные в виде пакетов, и стафилококки - скопления кокков, напоминающие грозди винограда.
Палочковидные бактерии - форма может быть в виде цилиндра, овоида различной длины и диаметра. Концы палочек закругленные, заостренные или резко обрубленные. Палочки, образующие споры, называются бациллами, не образующие споры - бактериями. Палочки, располагающиеся попарно, получили название диплобактерии, или диплобациллы, а располагающиеся в цепочку, - стрептобактерии, или стрептобациллы.
Извитые бактерии - это микроорганизмы, имеющие вид спирали. Они подразделяются на вибрионы, напоминающие слегка изогнутую запятую, спириллы, имеющие несколько крупных завитков, и спирохеты - бактерии с тонкими многочисленными завитками.
Строение бактериальной клетки . Ультраструктуру бактерий изучают с помощью электронно-микроскопических и микрохимических исследований, которые позволяют довольно точно определить структуру и составные части микробной клетки. Бактериальная клетка состоит из оболочки, цитоплазмы, ядерного вещества (рис.9).
Оболочка обладает значительной прочностью, упругостью, эластичностью, и благодаря этому создается как бы жесткий каркас микробной клетки, предохраняющий ее от неблагоприятных внешних воздействий и придающий ей постоянную форму (кокки, палочки). Оболочка имеет мельчайшие поры, она полупроницаема, через нее происходит обмен веществ с внешней средой.
Химический состав оболочки неоднороден: в ее составе обнаруживают азотистые и безазотистые соединения.
Оболочка бактерий представлена тремя структурами: внешним капсульным слоем, клеточной стенкой и цитоплазматической мембраной. Цитоплазма представляет собой дисперсную смесь коллоидов, состоящую из белков, воды, РНК (рибонуклеиновая кислота), липоидов, углеводов, минеральных веществ и др. Цитоплазма окружена тонкой цитоплазматической мембраной, состоящей из липопротеиновых и рибонуклеиновых компонентов. С цитоплазматической мембраной связаны ферментные системы, принимающие участие в обмене веществ с окружающей средой.
Цитоплазма содержит различные включения, наполненные клеточным соком, которые являются запасным питательным субстратом. В цитоплазме постоянно происходят процессы синтеза и распада веществ, т.е. осуществляются все функции, присущие живому организму.
Ядерное вещество бактериальной клетки, представленное ДНК (дезоксирибонуклеиновая кислота) в виде овальных и мелкозернистых включений, распределено в цитоплазме диффузно. Вокруг ДНК нуклеоида в цитоплазме бактерий расположены короткие двухцепочечные нити внехромосомной ДНК, получившие название плазмиды. Они управляют функцией устойчивости к лекарственным препаратам (R-плазмиды), выработки энтеротоксинов и обуславливают внехромосомную передачу наследственных свойств.
5) Колонии бактерий подсчитывают обычно через 2 суток, колонии грибов и дрожжей - через 5-7 суток. Колонии, как правило, подсчитывают с помощью лупы, не открывая чашек Петри. Для удобства отмечают просчитанную колонию точкой на наружной стороне дна чашки, пользуясь стеклографом или чернилами по стеклу. Колонии подсчитывают следующими способами: если они изолированы друг от друга, крупные и в небольшом количестве, то обычно их считают по всей поверхности чашки; при большом количестве выросших колоний дно чашки Петри делят на секторы (4-6-8 и т. д.). Подсчитывают в 2-3 секторах, находят, среднее арифметическое на один сектор, а затем умножают на количество секторов. Или подсчитывают количество колоний в каждом секторе и результаты суммируют; если колонии очень мелкие и их много, то следует пользоваться счетным аппаратом Вольфхюгеля. Аппарат состоит из черной: доски и стеклянной пластинки, разделенной на квадраты площадью 1 см2. Чашки необходимо поставить на доску вверх дном, покрыть стеклянной пластинкой и подсчитать колонии в квадратах по диагонали (10-12), затем рассчитать среднее арифметическое на один квадрат и пересчитать на площадь всей чашки, используя формулу S = πr2.
7) Роль микроорганизмов в круговороте веществ. Круговорот углерода. Роль бактерий в обмене углерода. Круговорот веществ — совокупность превращений химических элементов, из которых построены живые организмы. Основные факторы, определяющие доминирующую роль микробов в круговороте веществ, — широкое распространение микроорганизмов (например, слой плодородной почвы толщиной 15 см может содержать до 5 т микробной биомассы на гектар) и их необычайная метаболическая гибкость при высокой скорости обмена. Большое значение имеет узкая специализация отдельных групп микроорганизмов в отношении утилизируемых веществ. Поэтому некоторые этапы круговорота веществ могут осуществляться исключительно прокариотами. В природе все организмы разделяют на три группы. Продуценты — зелёные растения и микроорганизмы, синтезирующие органические вещества, используя энергию солнца, углекислый газ и воду. Потребители (консументы) — животные, расходующие значительную часть первичной биомассы на построение своего тела. Деструкторы — бактерии (в том числе актиномицеты) и грибы, разлагающие погибшие животные и растения; при этом органические вещества превращаются в неорганические, то есть происходит минерализация. Роль микроорганизмов в круговороте веществ. Круговорот углерода. Роль бактерий в обмене углерода. Круговорот углерода. Роль бактерий в обмене углерода. Взаимосвязь живых организмов на Земле особенно ярко выражена в круговороте углерода. Атмосферный воздух содержит около 0,03% С02, но продуктивность зеленых растений настолько велика, что весь запас углекислоты в атмосфере (2600-109 т С02) был бы истрачен за 20 лет — срок, ничтожно короткий в масштабах эволюции. Фотосинтез бы прекратился, если бы микроорганизмы, растения и животные не обеспечивали возвращение С02 в атмосферу в результате непрерывной минерализации органических веществ. Циклические превращения углерода и кислорода реализуются главным образом через два разнонаправленных процесса: кислородный фотосинтез и дыхание (либо горение в небиологических реакциях). • При кислородном фотосинтезе аэробные цианобактерии и зелёные растения основную часть окисленной формы углерода (СО2) переводят в восстановленное состояние, характерное для органических соединений (например, глюкозу), а восстановленную форму кислорода (Н20) окисляют до 02. Хотя анаэробные пурпурные и зелёные бактерии могут восстанавливать С02 до органических веществ, окисляя отличные от воды соединения (NH3, NO2, Н2, Fe2+, восстановленные соединения серы), вклад этих процессов в общую фиксацию СО, незначителен. В результате фотосинтетической фиксации С02 образуются сахара и другие соединения. Основная масса фиксированного углерода растений откладывается в виде полимерных углеводов (крахмал, целлюлоза). Поэтому сахара играют ведущую роль в питании всех живых организмов, нуждающихся в органической пище (организмы-гетеротрофы), и служат предпочтительными питательными веществами для большинства гетеротрофных микроорганизмов. • В присутствии кислорода полное окисление органических веществ до CO., осуществляют многие аэробные (псевдомонады, бациллы) и факультативно анаэробные (актиномицеты) бактерии, грибы, а также животные. В качестве примеров неполного окисления можно привести окисление Сахаров уксуснокислыми бактериями (Acetobacter, Gluconobacter) с образованием ацетата, образование лактата грибами порядка Mucorales (Rhizopus oryzae, R. nigricans и др.), образование глюконовой кислоты аспергиллами и пенициллами. Роль микроорганизмов в круговороте веществ. Круговорот углерода. Роль бактерий в обмене углерода. • В анаэробных условиях органические соединения расщепляются путём брожения (дрожжи, молочнокислые бактерии, пропионовокислые бактерии, бактерии семейства Entembacteriaceae), либо окисляются в процессе анаэробного дыхания при наличии акцепторов водорода. В роли акцепторов водорода выступают нитраты, сульфаты, карбонаты, фумараты, Fe3+: соответственно выделяют денитрифицирующие, сульфатредупирующие, метанобразующие бактериями. Метанобразующие бактерии (Methanobacterium, Melhanococcus, Methanosarcina) — строгие анаэробы, составляющие последнее звено анаэробной пищевой цепи. Выделяемый ими метан в аэробных условиях может окислятся до С02 метилотрофными бактериями (Methylomonas, Methytosinus, Melhylococcus). • Углерод извлекается из круговорота различными путями. Ионы карбоната, содержащиеся в морской воде, соединяются с растворёнными в ней ионами Са2+ и осаждаются в виде СаС03 (карбонат кальция). Последний также образуется биологическим путём в известковых структурах простейших, кораллов и моллюсков, откладываясь в качестве известняковых горных пород. Отложение неминерализованных органических остатков в условиях высокой влажности и недостатка кислорода приводит к накоплению гумуса, образованию торфа и каменного угля. Ещё один вид изъятия органического углерода из круговорота — отложения нефти, и газа (метана). • Деятельности человека неуклонно сдвигает баланс углерода в сторону образования С02. С одной стороны, это связано с интенсивным сжиганием нефти, угля и природного газа, а с другой — с уменьшением фотосинтетической фиксации углерода за счёт уничтожения лесных массивов, деградации почвы и загрязнения океана.
8) Круговорот азота. Роль бактерий в обмене азота. Азотфиксация. Денитрификация ( нитратное дыхание ). Ассимиляционная нитратредукция. Азот составляет 80% земной атмосферы; количество азота, участвующего в круговороте, исчисляется 108—109 т в год. Как газ азот химически инертен; он не может быть непосредственно использован растениями, животными и большинством микроорганизмов. Относительный дефицит связанного азота на поверхности Земли при практически неисчерпаемом его запасе в атмосфере подразумевает наличие определённого этапа, лимитирующего скорость круговорота. Этот этап — азотфиксация, осуществляемая исключительно азотфиксирующими бактериями. Промышленный синтез аммиака из азота и водорода составляет не более 5% фиксированного азота нашей планеты. Поэтому значение биологической азотфиксацин для жизни на планете огромно. Азотфиксация в природе осуществляется как свободноживущими микроорганизмами (несим-биотйческая азотфиксацин), так и бактериями, существующими в сообществе с растениями (симбиотическая азотфиксация). Несимбиотическая азотфиксация осуществляется бактериями рода Azotobacter, фиксирующими около 20 мг азота на 1 г использованного сахара, аноксигенными фототрофными бактериями, цианобактериями, кл остри днями, факультативными анаэробами Bacillus polymixa, Klebsiella pneumoniae, хемолитотрофными бактериями Alcaligenes latus, Xanthobacter autotrophicus, метил отрофны ми, метано генным и и сульфатредуцирующими бактериями). Круговорот азота. Роль бактерий в обмене азота. Азотфиксация. Денитрификация ( нитратное дыхание ). Ассимиляционная нитратредукция. Симбиотическая фиксация азота осуществляется бактериями рода Rhizobium (вызывают образование клубеньков у бобовых растений), актиномицетами рода Franckia (симбионты тропических растений), цианобактериями Anabaena azollae, Nostoc punctiforme. В отсутствие кислорода бактерии из нитрата, используемого в качестве акцептора водорода, образуют молекулярный азот. Этот процесс известен как денитрификация (диссимиляционная нитратредукция), или «нитратное дыхание», так как в данном случае роль нитрата в качестве окислителя аналогична роли молекулярного кислорода в аэробном дыхании. Способностью к денитрификации обладают многие факультативно аэробные бактерии (Pseudomonas aeruginosa, P. stutzeri, P. fluorescens, Bacillus Ucheniformis, Paracoccus denitrificans, Thiobacillus denitrificans). Ассимиляционная нитратредукция характерна для большинства микроорганизмов и для растений. Нитрат служит источником азота для построения клеточных компонентов. Источником азота для растений и микроорганизмов может быть и аммоний, усвояемый ими в процессе ассимиляции аммиака. • При разложении растительных и животных белков в почве освобождается аммоний. Собственно процесс аммонификации обусловлен деятельностью различных грибов и бактерий (Bacillus cereus, Proteus vulgaris, псевдомонады и др.). Если в почве достаточно кислорода, то аммоний подвергается нитрификации, которую осуществляют две труппы микроорганизмов, соответственно окисляя аммиак до нитрита (виды Nitrosomonas, Nitrosococcus, Nitrosospira, Nitrosolobus) и нитрит до нитрата {Nitrobacter, Nitrospina, Nitrococcus). Частично нитрификация осуществляется и при участии гетеротрофных бактерий (виды Arthrobacter), образующих нитрит из аммония, и грибов, способных окислять аммоний до нитрата. Однако скорость нитрификации у гетеротрофных бактерий в 103-104 раз меньше, чем у аутотрофных. • Нитрат — основное азотистое вещество почвы, используемое растениями в процессе роста. Практика удобрения почв навозом основана на способности микроорганизмов к минерализации органического азота. Нитраты легко выщелачиваются из почвы и, таким образом, часть связанного азота в виде солевых растворов удаляется с материков в океан. Связанный азот, необходимый для роста растений, удаляется из почвы также в процессе денитрификации, идущем с освобождением газообразного азота. Без денитрификации земной и атмосферный запас азота в конце концов сосредоточился бы в виде солей в океане, и жизнь стала бы возможной только на узкой прибрежной полосе. Удаление токсичных нитратов и мутагенных нитритов из пресной воды в реакциях денитрификации приводит к улучшению качества питьевой воды. Таким образом, микроорганизмы — естественные регуляторы количества связанного (то есть доступного для жизнедеятельности) азота в природе.
9) Превращение соединений фосфора. Фосфор имеет большое значение в жизнедеятельности организма. Без фосфора не могут синтезироваться белки, он в большом количестве входит в состав ядерного вещества и многих ферментов, участвует в так называемых реакциях фосфорелирования. Некоторые фосфорорганические компоненты – носители больших запасов энергии (нуклеиновые кислоты, липиды и др.).
В почве имеется много фосфора. По расчетам, его количество составляет 3 – 5 т/га. Особенно много этого элемента в черноземах, богатых гумусом (5 – 6 т/га). Фосфор в почве содержится в основном в органической, неусвояемой растением формой и в виде трудноусвояемых минеральных соединений. Органические соединения фосфора попадают в почву вместе с растительными остатками, а также с отмирающими микроорганизмами. Они представлены нуклеопротеидами, нуклеиновыми кислотами и т.д. Из разнообразия фосфорных микроорганизмов наибольший интерес представляют спорообразующие формы, так как они используются для приготовления бактериального удобрения фосфобактерина. Их относят к виду Bac. Megateriumvar. phosphatium. Это крупные палочки с закругленными концами, плотной оболочкой и зернистой цитоплазмой. Размеры клеток 5 – 6 мкм в длину и 1,8 – 2 мкм в ширину. В ранней стадии клетки расположены поодиночно и слабоподвижны, в дальнейшем они располагаются попарно или короткими цепочками и становятся неподвижными. При старении концы клеток приобретают конусообразную форму.
Клетки богаты органическими соединениями фосфора, нуклеопротеидами, образуют овальные эндоспоры, расположенные внутри клетки. Окрашиваются клетки по Граму, аэробы. Оптимальная температура роста 37°C. Колонии резкоокаймленные грязновато-белого цвета. Старые колонии вначале желтеют, а затем приобретают бурую окраску. На среде, содержащей фосфорорганические соединения и мел, вокруг колонии под влиянием кислот образуются зоны просветления. Фосфорные микробы энергично расщепляют органические соединения фосфора, освобождают фосфор в виде минеральных легкорастворимых солей фосфорной кислоты, доступных для растений. Чем больше таких микробов в почве, тем больше в ней доступного фосфора.
Превращение соединений серы. Сера содержится в организме животных и растений, входит в состав серосодержащих аминокислот (цистеин, цистин, метионин), витаминов группы B (биотин, тиамин), много ее в волосах и перьях. Органические соединения серы в почве представлены остатками животных и растений. Минерализация серы осуществляется микроорганизмами, которые в аэробных условиях доводят ее до сульфатов, а в анаэробных – восстанавливают серосодержащие белки до сероводорода и частично до меркаптанов.
Восстановленные соединения серы окисляют автотрофные (фотолитотрофы, хемолитотрофы) микробы. Среди них различают нитчатые, тионовые и фотосинтезирующие. Нитчатые хемолитотрофные серобактерии – аэробы и относятся к родам Beggiatoa, Theatric, Thioploca и другим.
Микроорганизмы, восстанавливающие соединения серы, – облигатные анаэробы. В таких условиях они в качестве конечного акцептора водорода используют сульфат. Донором водорода служат различные органические соединения и молекулярный водород. Процесс окисления органических соединений идет не до конца, основным продуктом бывает уксусная кислота, а побочным – сероводород. Образовавшийся газ может затем окисляться серобактериями, в результате чего накапливается биогенная сера.
Превращение соединений железа. Процесс окисления закисного железа в окисное осуществляется при участии железобактерий – окисляют комплексные соединения железа, а образующийся в результате гидроксид железа откладывается на поверхности клеток бактерий и придаёт клетке бурый цвет. Железобактерии чаще встречаются в болотах, ручьях, железистых источниках, озерах, дренажных трубах. 1. Наиболее важное значение из железобактерий имеют бактерии рода Leptothrix (L. ochraceae) – грамотрицательные прямые нитчатые палочки, в цепочках, окруженные трубчатым чехлом или свободно плавающие в виде отдельных клеток или коротких цепочек. Бактерии обитают в воде.
Свободные клетки подвижны за счет одного полярного жгутика. Чехлы бактерий пропитаны или покрыты окислами железа и марганца. Как правило, встречаются пустые чехлы без клеток бактерий. Скапливаясь, пустые чехлы образуют на дне водоемов охристые (жёлто-коричневый) осадки.
2. В окислении закисных солей железа принимают участие одноклеточные формы бактерии рода Gallionella (G. ferrugineae) – вибриоидные клетки со жгутиками. Клетки расположены на длинном плоском, спирально перекрученном стебельке. Одна сторона клетки вогнутая, другая – выпуклая. Из выпуклой стороны клетки выделяется наружу коллоидный гидроксид железа, из которого постепенно формируется стебелёк. При делении клетки стебелёк дихотомически ветвится. Стебелёк способен к самостоятельному росту и из него возникают новые клеточные образования.
10) Поступление веществ в клетку и выделение продуктов обмена в окружающую среду происходит у микроорганизмов через всю поверхность тела. Вещества питательной среды могут поступать в клетку только в растворенном состоянии. Клеточная стенка проницаема и задерживает лишь макромолекулы. Цитоплазматическая мембрана обладает полупроницаемостью. Она служит осмотическим барьером, проницаемость её для различных веществ неодинакова. Наиболее известны два пути проникновения веществ в клетку: осмос и адсорбция (специфический перенос). Активная роль в этих процессах принадлежит цитоплазматической мембране. Осмос представляет собой диффузию веществ в растворах через полупроницаемую перепонку (мембрану). Как известно, через такие мембраны могут диффундировать вещества, находящиеся в состоянии истинных растворов. Возникает осмос под действием разности осмотических давлений в растворах по обе стороны полупроницаемой мембраны. Таким образом, при осмотическом проникновении питательных веществ в клетку движущей силой служит разность осмотических давлений между средой и клеткой. Такой пассивный перенос веществ не требует затраты энергии и протекает до выравнивания концентрации с наружным раствором. Второй путь поступления веществ в клетку - активный - путём переноса их особыми, локализованными в цитоплазматической мембране веществами ферментной природы. Эти переносчики, называемые пермеазами, обладают субстратной специфичностью. Каждый транспортирует только определённое вещество, имеющее сходную с белком-переносчиком стереохимическую структуру молекулы. На внешней стороне цитоплазматической мембраны переносчик адсорбирует вещество - вступает с ним во временную связь и диффундирует комплексно через мембрану, отдавая на внутренней стороне её транспортируемое вещество в цитоплазму.
13) Важнейшими элементами являются органогены (углерод, водород, кислород, азот), которые используются для построения сложных органических веществ: белков, углеводов и липидов. Микроорганизмы содержат также зольные или минеральные элементы. Большая часть их химически связана с органическими веществами, остальные присутствуют в клетке в виде солей.
В количественном отношении самым значительным компонентом клетки является вода, которая составляет 75 - 85%; на долю сухого вещества, которое состоит из органических (белки, нуклеиновые кислоты, углеводы, липиды) и минеральных соединений, приходится 15-25%.
Вода. Все вещества поступают в клетку с водой, с ней же удаляются продукты обмена. Вода в микробной клетке находится в свободном состоянии как самостоятельное соединение, но большая часть ее связана с различными химическими компонентами клетки (белками, углеводами, липидами) и входит в состав клеточных структур.
Свободная вода принимает участие в химических реакциях, протекающих в клетке, является растворителем различных химических соединений, а также служит дисперсной средой для коллоидов. Содержание свободной воды в клетке может изменяться в зависимости от условий внешней среды, физиологического состояния клетки, ее возраста. Так, у споровых форм бактерий значительно меньше воды, чем у вегетативных клеток. Наибольшее количество воды отмечается у капсульных бактерий.
Белки (50-80% сухого вещества) определяют важнейшие биологические свойства микроорганизмов. Это простые белки - протеины и сложные - протеиды. Большое значение в жизнедеятельности клетки имеют нуклеопротеиды - соединение белка с нуклеиновыми кислотами (ДНК и РНК). Кроме нуклеопротеидов, в микробной клетке содержатся в незначительных количествах липопротеиды, гликопротеиды, хромопротеиды.
Белки распределены в цитоплазме, нуклеоиде, они входят в состав структуры клеточной стенки. К белкам принадлежат ферменты, многие токсины (яды микроорганизмов).
Видовая специфичность микроорганизмов зависит от количественного и качественного состава белковых веществ.
Нуклеиновые кислоты в микробной клетке выполняют те же функции, что и в клетках животного происхождения. ДНК содержится в ядре (нуклеоиде) и обусловливает генетические свойства микроорганизмов. РНК принимает участие в биосинтезе клеточных белков, содержится в ядре и цитоплазме. Общее количество нуклеиновых кислот колеблется от 10 до 30% сухого вещества микробной клетки и зависит от ее вида и возраста.
Углеводы (12-18% сухого вещества) используются микробной клеткой в качестве источника энергии и углерода. Из них состоят многие структурные компоненты клетки (клеточная оболочка, капсула и другие). Клетки микроорганизмов содержат простые (моно- и дисахариды) и высокомолекулярные (полисахариды) углеводы. У ряда бактерий могут быть включения, по химическому составу напоминающие гликоген и крахмал, они играют роль запасных питательных веществ в клетке. Углеводный состав различен у разных видов микроорганизмов и зависит от их возраста и условий развития.
Липиды (0,2-40% сухого вещества) являются необходимыми компонентами цитоплазматической мембраны и клеточной стенки, они участвуют в энергетическом обмене. В некоторых микробных клетках липиды выполняют роль запасных веществ.
Липиды состоят в основном из нейтральных жиров, жирных кислот, фосфолипидов. Общее количество их зависит от возраста и вида микроорганизма. Например, у микобактерий туберкулеза количество липидов достигает 40%, что обусловливает устойчивость этих бактерий к воздействию факторов внешней среды.
В клетках микроорганизмов липиды могут быть связаны с углеводами и белками, составляя сложный комплекс, определяющий токсические свойства микроорганизмов.
Минеральные вещества - фосфор, натрий, калий, магний, сера, железо, хлор и другие - в среднем составляют 2-14% сухого вещества.
Фосфор входит в состав нуклеиновых кислот, фосфолипидов, многих ферментов, а также АТФ (аденозинтрифосфорной кислоты), которая является аккумулятором энергии в клетке. Натрий участвует в поддержании осмотического давления в клетке. Железо содержится в дыхательных ферментах. Магний входит в состав рибонуклеата магния, который локализован на поверхности грамположительных бактерий.
Для развития микроорганизмов необходимы микроэлементы, содержащиеся в клетке в очень малых количествах. К ним относят кобальт, марганец, медь, хром, цинк, молибден и многие другие. Микроэлементы участвуют в синтезе некоторых ферментов и активируют их. Соотношение отдельных химических элементов в микробной клетке может колебаться в зависимости от вида микроорганизма, состава питательной среды, характера обмена и условий существования во внешней среде.
16) Влияние факторов внешней среды на микроорганизмы.
Действие на микроорганизмы физических факторов.
Температура – один из наиболее важных факторов в жизни микробов. Она может быть оптимальной, т.е. наиболее благоприятной для развития микробов;
Максимальной, когда подавляются жизненные процессы микробов;
Минимальной, ведущей к замедлению и прекращению роста.
Микроорганизмы, по чувствительности к температурным условиям объединены в 3 физиологические группы: психрофилы, мезофиллы и термофилы.
Психрофилы (криофильные) микроорганизмы – развиваются при низких температурах, обитатели холодных источников, глубоких озер и океанов. Например, светящиеся бактерии и железобактерии
Температурные границы: минимум -10 градусов, оптимум -8 - +5, максимум + 15 градусов.
Мезофильные микроорганизмы – развиваются при средних температурах 20 – 40 градусов. Большинство сапрофитов и все патогенные микроорганизмы. Температурные границ: минимум +20, оптимум 25 – 39 , максимум 40 градусов.
Термофильные микроорганизмы – развиваются при высоких температурах 40 – 80 градусов. Они встречаются в микрофлоре почвы, воды, теплых минеральных источниках, навозе, молоке и экскретах человека и животных. Температурные границы: минимум 28 – 35 градусов, оптимум 50-65, максимум 70 – 80 градусов.
Низкие температуры обычно не вызывают гибели микробов, а лишь задерживают их рост и размножение. Некоторые споры сохраняются жизнеспособность при температуре -253 градуса. Сальмонеллы длительно выживают во льду. Споры бацилл выдерживают температур +250 градусов в течение 3 суток. Низкие температуры приостанавливают гнилостные и бродильные процессы (на этом принципе основано использование холодильников, погребов для сохранения пищевых продуктов).
К высокой температуре особенно чувствительны вегетативные формы. На микробы эффективно действуют насыщенный водяной пар. Бактерицидное действие высоких температур вызывается повреждением рибосом, денатурацией белка и нарушением осмотического барьер. Применение высоких температур применяется для стерилизации.
Высушивание и вакуум приводят к обезвоживанию цитоплазмы и денатурации белков, действуют губительно на микроорганизмы. Более чувствительны к высушиванию вегетативные формы микробов. Споровые формы микробов в высушенном состоянии могут, сохранятся годами (bac. Anthraces, cl. Tetani), т. к. физиологическое состояние их не нарушается, количество свободной воды уменьшается до 40%, оболочка уплотняется, а жизненные функции полностью сохраняются (споры бацилл сибирской язвы в высушенном виде сохраняются несколько десятков лет).
Одним из методов консервирования пищевых продуктов является сублимация – обезвоживание при низкой температуре и высоком вакууме, которое сопровождается испарением воды, быстрым охлаждением и замораживанием (образовавшийся в продукте лед легко возгоняется, минуя жидкую фазу). Сублимационная сушка обеспечивает сохранение всех сахаров, витаминов, ферментов и др. компонентов. Высушивание в вакууме при низкой температуре не убивает бактерии и вирусы. Этот метод сохранения культур используется в производстве стабильных и с длительным сроком хранения живых вакцин (метод лиофилизации), витаминов, ферментов и др. биологических препаратов. Быстрое замораживание взвесей бактерий и вирусов при очень низкой температуре создает условия, при которых не происходит образования кристаллов и разрушения ими микроорганизмов.
Действие света (видимого излучения). Свет представляет собой электромагнитное излучение с длиной волны 400 – 780 нм. Естественным источником видимого излучения являются солнце, звезды, атмосферные разряды, люминесцирующие объекты и т.д. одни бактерии (пурпурные) хорошо переносят действие света, на других солнечный свет оказывает вредное влияние. Наиболее губительное действие оказывают прямые солнечные лучи. Различные виды облучения оказывают бактерицидное или бактериостатическое действие – затормаживает действие микробов, но не убивает их. Бактерицидность видимого излучения зависит от длины волны: чем она короче, тем в ней больше энергии.
Действие на микробы у/ф излучения – электромагнитные лучи с длиной волны 100 – 400 нм. Действуют на нуклеиновые кислоты. Микробицидное действие у/ф лучей основано на разрыве водородных связей и образовании в молекулах ДНК димеров тимина, приводящем к появлению нежизнеспособных мутантов. Они вызывают мутации, нарушают генетические процессы, инактивируют биосинтез жизненно важных компонентов клеток, что приводит к гибели микробной клетки.
Действие рентгеновского излучения – электромагнитное излучение с длиной волны 0, 005 – 2 мкм. К излучениям более чувствительны молодые клетки, находящиеся в стадии деления или роста. Более устойчивы Гр+ микробы, менее устойчивы Гр-.
Установлено, что тионовые бактерии, обитающие в залежах урановых руд, обладают высокой устойчивостью к радиоактивным излучениям. Микробов находили в воде атомных реакторов при концентрации ионизирующей радиации 2 – 3 млн.рад.
Ионизирующая радиация используется в практике стерилизации пищевых продуктов. Этот метод холодной стерилизации имеет ряд преимуществ перед тепловой: не изменяет качества продукта вследствие денатурации составных его частей (белки, полисахариды, витамины).
Гидростатическое давление на жизнедеятельность микроорганизмов. Микроорганизмы, устойчивее к высокому давлению, называются барофильными. Они существуют при давлении 100 МПа. Особенно выражена устойчивость у спор, которые сохраняются при давлении 200 МПа. Большинство микроорганизмов выдерживает давление до 65 МПа. Ингибирующее влияние высокого гидростатического давления взывает нарушение функции цитоплазматической мембраны, денатурацию белков, инактивацию ферментов.
Ультразвук – высокочастотные механические колебания упругой среды, не воспринимаемые ухом человека. Ультразвук, действуя на микроорганизмы, создает большую разницу в давлениях и повреждает клетку. Эффективность действия ультразвука понижается при содержании в среде протеина. Поэтому его использование для стерилизации молока не всегда дает желаемые результаты. В настоящее время ультразвук используется для стерилизации пищевых продуктов с низким содержанием белка, изготовления вакцин и дезинфекции предметов.
Аэронизация используется для обезвреживания воздуха цехов предприятий, жилых помещений, а также в медицинской и ветеринарной практике. Губительное действие оказывают отрицательно заряженные ионы. Положительно заряженные ионы задерживают рост бактерий лишь в больших концентрациях (106). Сила действия ионов зависит: от дозы – числа аэроионов на 1 см3 воздуха, длительности экспозиции; расстояния от источников ионов.
17) Действие на микроорганизмы химических факторов.
Химические вещества могут тормозить или полностью подавлять рост микроорганизмов. Действие химических веществ на микроорганизмы зависит от: природы вещества, физико-химического состава среды, концентрации, продолжительности контакта, температуры.
Спирты. Практически интерес представляет собой этиловый спирт. Его антимикробная активность проявляет способностью отнимать воду и свертывать белки. Бактерицидное действие проявляется уже с 20% - й концентрацией. Сильное противомикробное действие оказывает 70 градусный спирт. Высокие концентрации спирта (80 – 90%) в белковой среде образуют плотные белковые свертки, внутри которых могут, сохранятся живые бактерии.
Поверхностно-активные вещества обладают бактерицидным действием, к ним относятся жирные кислоты и мыла. Поверхностно-активные вещества накапливаются на поверхности клеточной стенки и вызывают резкое снижение поверхностного натяжения, что приводит к нарушению нормального функционирования клеточной стенки и цитоплазматической мембраны.
Фенол, крезол и их производные (карболовая кислота) первоначально повреждают клеточную стенку, а затем и белки клетки.
Красители - задерживают рост бактерий. Они действуют медленно, но более избирательно. К ним относятся бриллиантовый зеленый, метиленовая синь.
Окислители – к ним относятся перманганат калия и перекись водорода. Выделяя атомарный кислород, вызывают цепную реакцию свободно-радикального перекисного окисления липидов, что ведет к деструкции мембран и белков микроорганизмов.
Соли тяжелых металлов (свинец, цинк, серебро, ртуть) вызывают денатурацию белков клетки. Ряд металлов обладает олигодинамическим действием (бактерицидным). К таким относятся: серебро, золото, цинк, олово, свинец. Например, посуда из серебра, посеребренные предметы при контакте с водой сообщают ей бактерицидные свойства по отношению ко многим видам бактерий.
Механизм олигодинамического действия заключается в том, что положительно заряженные ионы металлов адсорбируются отрицательно заряженной поверхностью бактерий и изменяют проницаемость клеточной стенки. При этом происходит нарушение питания и размножения бактерий. К солям тяжелых металлов также очень чувствительны и вирусы.
Формальдегид – применяется 40% раствора формалина. Его противомикробное действие вызвано тем, что он, вступая в реакцию с белками, вызывает их денатурацию. Убивает как вегетативные формы бактерий, так и споры. Применяется для обезвреживания столбнячного токсина, благодаря чему они превращаются в анатоксины.
По устойчивости к действию поваренной соли различают:
- негалофильные микроорганизмы – развиваются при содержании поваренной соли менее 0,01%
- слабогалофильные (толерантные) лучше всего растут в среде с содержанием поваренной соли 1 – 5%
- среднегалофильные (умеренные) переносят 5 – 20% поваренной соли
- сильногалофильные (экстремальные) микроорганизмы развиваются при концентрации поваренной соли от 12 – 15% до насыщенных растворов.
18,19) Влияние на микроорганизмы биологических факторов.
Между микробами в природе постоянно происходит борьба за существование.
Живые организмы объединены в устойчивые экологические системы – биоценозы. Для каждого из них характерны видовое и количественное соотношения популяций, структура, взаимоотношения и другие признаки.
Между различными группами микробов существуют разнообразные взаимоотношения. Они могут проявляться в форме симбиоза, метабиоза, сателлизма, синергизма, вирогенизма, антагонизма, комменсализма и т.д.
Симбиоз – сожительство организмов разных видов, приносящее им взаимную пользу. Они совместно развиваются лучше, чем каждый из них в отдельности. Иногда приспособленность организмов друг к другу становиться очень глубоким, и они утрачивают способность существовать порознь.
- симбиоз гриба и сине-зеленой водоросли – лишайники. Гриб обеспечивает водоросли прикрепление и защиту, а также снабжение водой и неорганическими солями. Водоросль предоставляет грибу продукты фотосинтеза.
- симбиоз клубеньковых бактерий и бобовых растений. Клубеньковые азотфиксирующие бактерии, обеспечивая азотом растения, стимулируют их рост, деление клеток и образование клубеньков. От растений бактерии получают углеводы и минеральные соли.
- симбиоз аэробов и анаэробов в замкнутой системе.
Метабиоз – вид взаимоотношений, когда один организм продолжает процесс вызванный другим, освобождая его от продуктов жизнедеятельности и теми самым создавая условия для его дальнейшего развития, т.е. продукты жизнедеятельности одного вида служат источником питания другого вида. Например: нитрифицирующие и аммонифицирующие бактерии. Нитрифицирующие бактерии используют для метаболизма аммиак – продукт жизнедеятельность аммонифицирующих почвенный бактерий.
Сателлизм – стимуляция роста и развития одних микроорганизмов за счет продуктов жизнедеятельности других.
Так, дрожжи – продуценты витаминов – способствуют нормальной жизнедеятельности других микроорганизмов, нуждающихся в витаминах, но неспособных их синтезировать в достаточном количестве или вообще не способны к такому синтезу.
Синергизм – характеризуется усилением физиологических функций у членов микробной ассоциации (дрожжи и молочнокислые бактерии в образовании молочнокислых продуктов).
Вирогенизм – совместное сосуществование некоторых бактерий, дрожжей и простейших с вирусами.
Паразитизм – это такое отношение между микробами, когда пользу от сожительства получает лишь паразит, нанося вред хозяину, что обычно приводит к гибели последнего.
Антагонизм – борьба микроорганизмов за кислород, пищевые веществ и место обитания. Определенные виды, которые приспособились к данной среде, обладают антагонизмом по отношению в видам, попадающим в новую среду.
Например, синегнойная палочка подавляет рост сальмонелл, бацилл сибирской язвы, стафилококков и др.
Антагонизм возможен внутри одного и того же вида. Антагонистические отношения возникают в результате недостатка питательных веществ и тогда одни микробы вынуждены питаться за счет других.
Некоторые бактерии, грибы и высшие растения вырабатывают различные вещества:
Антибиотики – вещества, губительно действующие на другие микробы
- бактериофаги – вирусы бактерий
- фитонциды – летучие вещества многих растений
Комменсализм – неярко выраженная форма сожительства микробов с другими организмами, при этом один организм использует пищи ил выделения другого, не принося ему вреда.
Комменсалы – представители нормальной микрофлоры животных, обитающие в ЖКТ, дыхательных путях, на коже, а также эпифитные микробы растений. 20,22) Модификацией называют ненаследственную изменчивость, которая возникает под воздействием факторов внешней среды и, как правило, не оказывает влияния на развитие последующих поколений, т.е. модификация не передается по наследству и утрачивается при восстановлении обычных для микроорганизма условий. Например, добавление к питательной среде солей лимонной кислоты увеличивает интенсивность ароматообразования у гетероферментативных молочнокислых стрептококков; недостаток кальция в среде вызывает повышение спорообразования и слизистый рост у бацилл сибирской язвы; неорганическое железо стимулирует образование токсинов микроорганизмами; глицерин и аланин обусловливают полиморфизм у холерных вибрионов и др. Микробы могут также временно изменять свою ферментативную способность.
Диапазоны модификационной изменчивости ограничиваются совокупностью фенотипов, которые создаются на основе определенного генотипа.
Изменения, появляющиеся в результате модификации, могут сохраняться в течение нескольких поколений.
При модификации не нарушается нормальное равновесие физиологических процессов в клетке и соответствие между клеткой и средой.
В связи с этим модификации также называют адаптивными реакциями на внешние раздражения, которые регулярно повторяются в нормальных условиях жизни.
Основы изменчивости бактерий: Модификации - фенотипические изменения какого-либо признака или нескольких признаков микроорганизмов. Модификации проявляются в изменении морфологических, биохимических и других признаков с последующей их реверсией к первоначальному фенотипу после устранения действия фактора, вызвавшего их образование.
Мутации - изменение первичной структуры ДНК, проявляющееся наследственно закрепленной утратой или изменением какого-либо признака или группы признаков. Факторы, вызывающие мутации, известны как мутагены. К появлению спонтанных мутаций приводят ошибки репликации, неправильное формирование пар оснований или структурные искажения ДНК под действием естественных мутагенов. Индуцированные мутации получают под влиянием каких-либо мутагенов (химические вещества, излучение, температура и др.) в эксперименте. По количеству мутировавших генов различают генные и хромосомные мутации.
Генетические рекомбинации. Бактерии способны обмениваться генетическим материалом и, по аналогии с половым размножением, давать начало потомству с новыми свойствами. При этом образуется ДНК, которая содержит гены обеих родительских клеток. Такую ДНК называют рекомбинантной. У потомства, или рекомбинантов, наблюдается заметное разнообразие признаков, вызванное смешением генов. Известны три способа получения рекомбинантов: путем трансформации, трансдукции и конъюгации.
21)Рекомбинационная изменчивость у бактерий.
Рекомбинация генов – процесс образования смешанного потомства в результате генетического обмена между двумя клетками. Вследствие рекомбинации образуются новые бактериальные клетки (рекомбинанты) с признаками обеих клеток. Рекомбинанты содержат основной набор генов реципиента и определенную часть клеток донора. У бактерий к образованию рекомбинантных хромосом ведет три способа переноса генетической информации: трансформация, конъюгация, трансдукция.
Трансформация – это изменение свойств бактериальной клетки в результате переноса информации, при котором фрагмент ДНК клетки-донора проникает в клетку – реципиент родственного штамма или вида. Впервые явление трансформации у бактерий наблюдал Ф.Гриффитс в 1928 г. В 1944 г. О.Эвери , К.Мак-Леод и М.Мак-Карти экспериментально доказали, что трансформирующим веществом является ДНК.
Не все клетки бактерий способны воспринимать ДНК. Клетки, которые воспринимают трансформирующую ДНК, называются компетентными. Они синтезируют специфический белок. Состояние компетентности связано со стадией активного размножения бактерий и появляется в начале логарифмической фазы. Процесс трансформации происходит в несколько стадий: 1) адсорбция и восприятие бактериями-реципиентами фрагмента двунитчатой ДНК клетки-донора (обычно не более 5 генов); 2) эклипс– скрытый период, характеризующийся отсутствием биологического действия ДНК.В этот период двунитчатая ДНК распадается: одна цепь разрушается, другая – участвует в последующих стадиях; 3) интеграция(включение) трансформирующей ДНК в хромосому реципиента в результате разрыва и воссоединения хромосомы донора; 4) экспрессия (проявление генов), т.е. размножение трансформированного клона клеток, потомство которых будет иметь измененный ген. Таким образом, при трансформации происходит замещение одного генетического участка другим. При этом процессе происходит трансформация чаще одного какого-нибудь признака, реже – двух, так как в реципиентную клетку проникает малый фрагмент ДНК. Различают гомотрансформацию – перенос генетической информации от одного штамма к другому в пределах одного вида, и гетеротрансформацию – перенос ДНК от одного вида к другому.
Конъюгация – передача генетического материала из клетки в клетку при непосредственном контакте путем образования цитоплазматического мостика между бактериями. Явление конъюгации впервые описали в 1946 г. Дж.Ледерберг и Э.Татум. Способность бактериальной клетки конъюгировать связана с наличием в ней полового фактора F – внехромосомой автономной детерминанты (F-плазмиды). Доноры содержат половой фактор (F+ ), реципиенты – не содержат (F-). Доноры способны синтезировать дополнительные поверхностные структуры – F-пили. Эти поверхностные структуры имеют форму ворсинок. Количество F-пилей соответствует числу копий полового фактора. Половые ворсинки представляют собой полые цилиндрические отростки, образованные белком пилином. Формируются F-пили только у активно растущей клетки за 4-5 мин и в течение такого же промежутка времени сохраняются на ее поверхности, затем сбрасываются. Процесс конъюгации начинается с прикрепления клетки-реципиента к кончику F-пиля, затем клетки приближаются друг к другу. Образуется конъюгационный цитоплазматический мостик, через который и происходит передача донорского генетического материала. Конъюгация бывает внутривидовая (между штаммами одного вида), межвидовая, межродовая.
Трансдукция – это перенос генетического материала от одних бактериальных клеток к другим с помощью фага. Трансдуцирующий фаг переносит фрагмент ДНК от клетки донора к реципиенту и вводит эту ДНК таким же образом, как и свою собственную ДНК.
Общая (неспецифическая, генерализованная, множественная) трансдукция - перенос любого гена или одновременно нескольких генов. Трандуцирующий фаг выступает в качестве "пассивного" переносчика генетического материала бактерии, захватывая фрагмент бактериальной ДНК место своего генома. Включения фаговой ДНК в бактериальную хромосому не происходит. Такие трансдуцирующие фаги получили название дефектных фагов. Специфическая (ограниченная) трансдукция характеризуется способностью фага переносить определенные гены. При взаимодействии таких фагов с клетками реципиента происходит включение гена клетки донора в хромосому реципиента вместе с ДНК дефектного фага, т.е. происходит лизогенизация бактерий-реципиентов. Бактерии, лизогенизированные дефектным фагом, невосприимчивы к последующему заражению тем же вирулентным фагом. Вместе с тем эти бактерии не способны в обычных условиях образовывать зрелый фаг.Абортивная трансдукция – перенесенный фагом фрагмент хромосомы донора на включается в хромосому клетки-реципиента, а располагается в ее цитоплазме и может в таком виде функционировать. Во время деления клетки трансдуцированный фрагмент ДНК донора может передаваться только одной из двух дочерних клеток, т.е. наследуется однолинейно и в конечном итоге утрачивается в потомстве.
Таким образом, все три процесса генетической рекомбинации у бактерий заключаются в переносе фрагмента ДНК от одной клетки к другой. При трансформации в бактерию-реципиент входит свободная ДНК; в процессе конъюгации происходит перенос фрагмента ДНК при образовании цитоплазматического мостика между бактериями; при трансдукции фаг захватывает участок хромосомы бактерии-донора и передает реципиенту; причем, при всех трех процессах переносится не вся хромосома, а ее фрагмент.
23) Иммунология – изучает закономерности проявления, механизмы и способы управления иммунитетом, антигены и антитела, вопросы аллергии, диагностики и специфической профилактики инфекционных болезней.
24,25) Инфекция и иммунитет.
Инфекция и инфекционный процесс.
Инфекция – это комплекс биологических процессов, которые возникают в результате проникновения патогенных микроорганизмов в макроорганизм.
Формы инфекции:
- септицемия – микробы размножаются в крови, и проникают во все органы и такни (сибирская язва).
- бактериемия – кровь переносит микробы, а размножение происходит в других тканях (бруцеллез, туберкулез)
- токсемия – микробы, размножаются в поврежденной ткани, образуя токсины, попадающие в кровоток (столбняк, ботулизм).
Инфекционный процесс – внедрение в восприимчивый организм, в определенных условиях окружающей среды, патогенных микробов, вызывающих нарушение постоянства внутренней среды и физиологических функций всего организма животного.
Крайней степенью инфекционного процесса является инфекционная болезнь.
Инфекционные болезни отличаются от других болезней следующими признаками:
- наличием живого возбудителя – т.е. микроорганизма
- специфической реакцией организма на конкретный возбудитель
- приобретение иммунитета к данному микробу (невосприимчивости к повторному заражению)
- заразностью, т.е. микробоносительством – состояния, при котором после выздоровления, в организм длительное время сохраняются микробы
Для возникновения и развития инфекционного заболевания необходимо:
- наличие патогенного микроба
- восприимчивый макроорганизм
- определенные условия внешней среды, в котором происходит взаимодействие между микроорганизмом и макроорганизмом.
Классификация инфекций по количеству возбудителей: простые или моноифекции – вызываются одним видом возбудителя и смешанные или ассоциативные – вызываются двумя или несколькими видами микробов.
Классификация инфекций по последовательности включения в инфекционный процесс возбудителя:
- суперинфекции – заболевание возобновляется до выздоровления больного организма
- реинфекции – организм заражается повторно после перенесенного заболевания
- рецидив – организм заболевает повторно за счет оставшихся в организме возбудителей
- вторичные (секундарные) инфекции – в уже развившийся процесс включается новый возбудитель.
Инфекции подразделяются на: экзогенные – возбудитель поступает из окружающей среды с пищей, водой, воздухом, почвой и т.д. и эндогенные – вызываются представителями нормальной микрофлоры (условно патогенные микроорганизмы), они постоянно находятся в организме.
Динамика развития инфекционного процесса:
- развитие инфекционного процесса складывается из следующих периодов:
А) инкубационного
Б) продромального
В) разгара болезни
Г) угасания болезни
Д) выздоровления (реконвалесценция)
Инкубационный период – это период с момента внедрения патогенного микроба и до появления первых признаков болезни.
Колеблется от нескольких часов (токсоинфекции) до нескольких месяцев и лет.
Продолжительность инкубационного периода зависит от степени общей резистентности и специфического иммунитета организма, его реактивности, влияния вредных факторов внешней среды, условий кормления и содержания, дозы и вирулентности возбудителя.
В этот период происходят размножение и накопление микробов и их токсинов, суммация образующихся раздражений, повышение реактивности организма к возбудителю и его токсинам.
Продромальный период – это период предвестников болезни. Наступают характерные для данной болезни симптомы и развиваются неспецифические, общие для многих болезней признаки (слабость, потеря аппетита, отказ от корма, температура).
Период разгара болезни – период основных проявлений болезни. Типичными признаками болезни являются: лихорадка, воспаление, поражение нервной системы. Наблюдаются нарушения со стороны ССС (сердечно-сосудистой системы), органов дыхания, пищеварения, мочевыделения и др.
Период угасания болезни – наступает при благоприятном исходе болезни. В одних случаях болезнь заканчивается кризисом – быстрым понижением температуры, сопровождающимся потоотделением.
В других случаях выздоровление характеризуется лизисом – постепенным понижением температуры и ослаблением явлений болезни.
Период выздоровления (реконвалесценция) – завершающий этап инфекционной болезни. Продолжительность периода колеблется от нескольких дней до нескольких недель.
Клиническое выздоровление при многих инфекционных болезнях не совпадает по времени с освобождением организма от возбудителя. В некоторых случаях развивается обострение болезни, рецидивы, осложнения, вторичные инфекции.
Патогенность и вирулентность. Факторы вирулентности.
Патогенность – это потенциальная способность определенных видов микробов вызывать инфекционное заболевание.
Патогенные микробы характеризуются специфичностью действия, т.е. каждый вид способен вызывать определенную инфекционную болезнь.
Микроб не вызывает заболевание если он не обладает вирулентностью.
Вирулентность – степень болезнетворности возбудителя.
Вирулентность – величина непостоянная, она изменяется под влиянием многих факторов. Ее можно повысить последовательными заражениями восприимчивых лабораторных животных и ослабить при культивировании в неблагоприятных условиях.
За единицу вирулентности принята минимальная смертельная доза DLM – это наименьшее количество микробных клеток, способное вызывать гибель восприимчивых животных. При изучении вирулентности определяют среднюю летальную дозу (LD50), при которой погибает 50% подопытных животных.
Факторы вирулентности:
- адгезия
- инвазивность
-токсинообразование
-капсулообразование
Адгезия. Адгезивность обуславливается специальными приспособлениями (фимбрии, адгезины, лиганды), расположенными на поверхности микробов, а также наличием соответствующих рецепторов на чувствительных клетках макроорганизма.
Адгезивную функцию выполняют специальные антигены – белки-лектины, которые содержатся в фимбриях патогенных штаммов микробов.
Штаммы микробов, лишенные таких антигенов, не способны прикрепляться к клеткам определенных органов.
Инвазивность (инвазионность) – соединительная ткань содержит гиалуроновую кислоту, обладающую способностью сопротивляться проникновению в ткань различных посторонних веществ, в том числе и патогенных микробов. Этот защитный барьер может быть разрушен ядовитыми веществами микробов, которые получили название «факторы распространения или проницаемости». Они характеризуются высокой активностью, действуют в очень малых дозах, разрушаются при температуре 60 градусов в течение 30 минут, обладают ферментативными свойствами, усиливают местное первичное действие патогенных микробов, поражающих соединительную ткань и способствуют развитию общей инфекции.
Токсинообразование. Различают экзотоксины и эндотоксины.
Экзотоксины обладают выраженным специфически токсическим действием. Характеризуются резко выраженной токсичностью, действуют на восприимчивый организм в малых дозах. По химическому составу принадлежат к веществам белковой природы. Чувствительны к действию света, кислорода и температуры. При температуре 60 – 80 градусах разрушаются в течение 10 минут, при кипячении – моментально. Одни экзотоксины (столбнячный) разрушаются под влиянием пищеварительных ферментов, поэтому являются безвредными при введении через рот; другие (ботулинистический, патогенные стафилококки) не разрушаются в желудке и кишечнике и вызывают отравление организма при пероральном введении.
Эндотоксины – прочно связаны с телом бактериальной клетки, менее токсичны, поражают в больших дозах и выделяются во внешнюю среду только после разрушения микробной клетки. Эндотоксины отличаются от экзотоксинов высокой устойчивостью к температуре, выдерживают кипячение и стерилизацию при 120 градусах до 30 минут. Эндотоксины обнаружены у возбудителей колибактериоза, сальмонеллеза, туберкулеза, бруцеллеза.
Капсулообразование – обеспечивает устойчивость микробов к фагоцитозу и антителам, увеличивает их инвазионные свойства. Вирулентность капсульных микробов связана с токсическими веществами, находящимися в капсуле.
По химическому составу капсульное вещество состоит у одних видов микробов из сложных полисахаридов, у других – из протеинов. Оно может быть неодинаковым у отдельных штаммов одного и тоже вида и может быть сходным у разных бактерий.
27,28) центральный и периферические органы иммунитета. Клетки участвующие в иммунных реакциях.
«Виды иммунитета» Таблица
Вид иммунитета |
Способ проявления |
Врожденный (естественный) |
Сопротивляемость к заболеваниям с рождения |
Приобретенный (естественный) |
Формирование антител после инфекционной болезни |
Активный (искусственный) |
Возникает после прививки |
Пассивный (искусственный) |
Появляется в результате введения сыворотки |
Иммунитет – это способ защиты организма от живых тел и существ, несущих на себе признаки генетической чужеродности.
1)Специфический. 1.1)Искусственный активный:
Вакцины – это ослабленные или убитые микробы.
Ослабленные вакцины – это живые вакцины. На живые вакцины организм вырабатывает более напряженный иммунитет, он более длительный, т.е. живые вакцины более эффективны, чем ослабленные.
Инактивированные вакцины – это вакцины убитые воздействием температуры или химических веществ.
Иммунитет на инактивированные вакцины менее напряженный, менее продолжительный.
Антиген и антител взаимодействуют между собой строго специфично.
1.2)Искусственный пассивный иммунитет это введение иммунных сывороток, т.е. введение готовых антител. Иммунные сыворотки получают путем гипериммунизации (многократной иммунизации) животных, ослабленными возбудителями. Биофабрики занимаются получением иммунных сывороток.
2.1)Естественный пассивный иммунитет: плацентарный иммунитет, который передается через плаценту плоду.
Колостральный иммунитет передается через молозиво новорожденному.
2.2)Естественный активный иммунитет: возникает после естественного переболевания. Стерильный – память об антигене остается в Т-клетках памяти.
Не стерильный – с исчезновением возбудителей из организма практически исчезает невосприимчивость.
29)Иммунный ответ – это цепь последовательных сложных кооперативных процессов, идущих в иммунной системе в ответ на действие антигена в организме.
Различают:
1) первичный иммунный ответ (возникает при первой встрече с антигеном);
2) вторичный иммунный ответ (возникает при повторной встрече с антигеном).
Любой иммунный ответ состоит из двух фаз:
1) индуктивной; представление и распознавание антигена. Возникает сложная кооперация клеток с последующей пролиферацией и дифференцировкой;
2) продуктивной; обнаруживаются продукты иммунного ответа.
При первичном иммунном ответе, индуктивная фаза может длиться неделю, при вторичном – до 3 дней за счет клеток памяти.
В иммунном ответе антигены, попавшие в организм, взаимодействуют с антигенпредставляющими клетками (макрофагами), которые экспрессируют антигенные детерминанты на поверхности клетки и доставляют информацию об антигене в периферические органы иммунной системы, где происходит стимуляция Т-хелперов.
Далее иммунный ответ возможен в виде одного из трех вариантов:
1) клеточный иммунный ответ;
2) гуморальный иммунный ответ;
3) иммунологическая толерантность.
Клеточный иммунный ответ – это функция T-лимфоцитов. Происходит образование эффекторных клеток – T-киллеров, способных уничтожать клетки, имеющие антигенную структуру путем прямой цитотоксичности и путем синтеза лимфокинов, которые участвуют в процессах взаимодействия клеток (макрофагов, T-клеток, B-клеток) при иммунном ответе. В регуляции иммунного ответа участвуют два подтипа T-клеток: T-хелперы усиливают иммунный ответ, T-супрессоры оказывают противоположное влияние.
Гуморальный иммунитет – это функция B-клеток. Т-хелперы, получившие антигенную информацию, передают ее В-лимфоцитам. В-лимфоциты формируют клон антителопродуцирующих клеток. При этом происходит преобразование B-клеток в плазматические клетки, секретирующие иммуноглобулины (антитела), которые имеют специфическую активность против внедрившегося антигена.
Образующиеся антитела вступают во взаимодействие с антигеном с образованием комплекса АГ – АТ, который запускает в действие неспецифические механизмы защитной реакции. Эти комплексы активируют систему комплемента. Взаимодействие комплекса АГ – АТ с тучными клетками приводит к дегрануляции и выделению медиаторов воспаления – гистамина и серотонина.
При низкой дозе антигена развивается иммунологическая толерантность. При этом антиген распознается, но в результате этого не происходит ни продукции клеток, ни развития гуморального иммунного ответа.
Иммунный ответ характеризуется:
1) специфичностью (реактивность направлена только на определенный агент, который называется антигеном);
2) потенцированием (способностью производить усиленный ответ при постоянном поступлении в организм одного и того же антигена);
3) иммунологической памятью (способностью распознавать и производить усиленный ответ против того же самого антигена при повторном его попадании в организм, даже если первое и последующие попадания происходят через большие промежутки времени).
30)Аллергия, антигены и антитела. Аллергия - это форма иммунного ответа. Цель - удаление чужого, механизм - приведение в действие воспаления (III- стадия аллергических реакций, патофизиологическая).
Термин "аллергия" предложил в 1906 г. Пирке. Хорошо известна туберкулиновая реакция его имени. Он писал: "Вакцинированный относится к вакцине, сифилитик - к вирусу сифилиса, туберкулезный - к туберкулину, получивший сыворотку - к последней иначе, чем индивидуум, не встречавшийся с этими агентами. Он, однако, очень далек от состояния нечувствительности. Все, что мы можем о нем сказать - это его реактивность является измененной. Для этого общего понятия измененной реакции я предлагаю выражение "аллергия". Т.е. аллергическая реакция - повышенная чувствительность организма, возникает после повторного введения антигена (аллергена).
Определенные вещества, вызывающие у человека и животных состояние повышенной чувствительности (гиперчувствительности - ГЧ), т.е. аллергию, называются аллергенами. Состояние аллергии может вызвать повторное введение генетически чужеродного вещества.