

Билет № 1
Биоразнообразие. Краткая характеристика. Прокариоты и эвкариоты. Основные типы питания. Значение и вопросы сохранения биоразнообразия.
Биологическое разнообразие на Земле – это около 2 млн известных и порядка 10 млн реально существующих, но пока еще не описанных наукой видов.
Теоретическая скорость исчезновения видов должна составлять 4 вида в год. Сегодня скорость исчезновения видов превышает естественный ход эволюции в среднем в 5000 раз. Средняя продолжительность существования вида – около 4 млн. лет, а на Земле существует по разным оценкам до 10-12 млн. видов. С такой скоростью исчезновения видов весьма вероятно, что половина видов наземных организмов может исчезнуть в ближайшие 50 лет…
Cейчас происходит самое значительное за последние 65 млн. лет исчезновение видов растений и животных, наблюдается деградация и гибель многих ценных ресурсных сообществ в первую очередь ропических лесов, в которых на площади в 1 га можно встретить до 200 видов только древесных растений, не считая тысяч видов беспозвоночных, нескольких десятков птиц и других многочисленных животных;
прибрежных коралловых рифов с огромным многообразием водных беспозвоночных и сотнями видов рыб;
в умеренной зоне распахиваются степи;
повсеместно загрязняются реки и воды мирового океана.
Проблемы изучения, сохранения и рационального использования природных ресурсов (включая растительный и животный мир) в современных условиях имеют не только научную актуальность, но и напрямую связаны с вопросами политики и экономики. Это в особой степени проявляется в подписании более 100 странами мира в 1992 г. в Рио-де-Жанейро Конвенции о биологическом разнообразии. Осознавая вечную ценность биоразнообразия как необходимого условия устойчивого функционирования биосферы, а также его широкое ресурсное, экологическое, научное, социально-экономическое, культурно-воспитательное, рекреационное и эстетическое значение, Республика Беларусь также подписала, а в июне 1993 г. ратифицировала Конвенцию о биологическом разнообразии.
Прокариоты (от др.-греч. pro – «перед», «до» и karyon – «ядро») или безъядерные – одноклеточные живые организмы, не обладающие (в отличие от эукариот) оформленным клеточным ядром. Для клеток прокариот характерно отсутствие ядерной оболочки, ДНК упакована без участия гистонов. К прокариотам относятся царства бактерии и археи. Генетический материал прокариот представлен одной молекулой ДНК, замкнутой в кольцо, т.е. имеется только один репликон. В клетках также отсутствуют органоиды (называемые митохондриями и хлоропластами), имеющие мембранное строение
Эукариоты или ядерные (лат. Eukaryota от др.-греч. eu – «хорошо» и karyon – «ядро») – надцарство живых организмов, клетки которых содержат ядра. Эукариотическими организмами являются все животные, растения, грибы, а также группы организмов под общим названием протисты. Они могут быть одноклеточными и многоклеточными, но все имеют общий план строения клеток. Считается, что все эти столь несхожие организмы имеют общее происхождение, поэтому группа ядерных рассматривается как монофилетический таксон наивысшего ранга. Согласно наиболее распространенным гипотезам, эукариоты появились 1,5–2 млрд. лет назад. Важную роль в эволюции эукариот сыграл симбиогенез – симбиоз между эукариотической клеткой, видимо, уже имевшей ядро и способной к фагоцитозу, и проглоченными этой клеткой бактериями –предшественниками митохондрий и хлоропластов.
Эукариотические клетки в среднем намного крупнее прокариотических, причем разница в объеме достигает тысяч раз. Клетки эукариот включают около десятка видов различных структур, известных как органоиды (или органеллы), из которых многие отделены от цитоплазмы одной или несколькими мембранами. В прокариотических клетках всегда присутствуют клеточная мембрана, рибосомы (существенно отличные от эукариотических рибосом) и генетический материал – бактериальная хромосома, или генофор, однако внутренние органоиды, окруженные мембраной, встречаются редко. Ядро – это часть клетки, окруженная у эукариот двойной мембраной (двумя элементарными мембранами) и содержащая генетический материал, а именно молекулы ДНК, «упакованные» в хромосомы. Ядро обычно одно, но бывают и многоядерные клетки.
Альфа-, бета- и гамма разнообразие. Биологическое разнообразие - вариабельность живых организмов из всех источников, включая, наземные, морские и иные водные экосистемы и экологические комплексы, частью которых они являются. Биологическое разнообразие включает в себя разнообразие в рамках вида, между видами и разнообразие экосистем.
Альфа-разнообразие - характеристика разнообразия внутри местообитания или внутри сообщества на видовом уровне.
Бета-разнообразие - показатель, измеряющий степень дифференцированности видов по градиентам местообитаний.
Гамма-разнообразие - показатель разнообразия на территориальном уровне, объединяющий альфа- и бета-разнообразие.
Сформулированные немецким гидробиологом Августом Тинеманом в 1939 г. законы экологического разнообразия гласят: чем разнообразнее условия существования в пределах биотопов (больше размерность экологической ниши), тем больше число видов в данном биоценозе.
Научный (академический) интерес к проблеме биоразнообразия связан с возможностью познания механизмов формирования структуры сообществ и экосистем разного масштаба. Как подчеркивает Р. Уиттекер (1980), сообщества «...являются функциональными системами дифференцированных по нишам видов, а структура сообщества, дифференцированная во времени и пространстве, значимость и разнообразие видов – это взаимосвязанные проявления организации видов в сообществах».
Прикладной (природоохранный) интерес к проблеме биоразнообразия связан со следующими причинами:
нарушение экосистемных и биосферных функций:
обеспечение оптимального газового состава атмосферы,
биологическая очистка от загрязняющих веществ,
сохранение способности экосистем преобразовывать солнечную энергию, сохранение плодородия почв и др.;
морально-эстетические факторы («Мы в ответе за тех, кого приручили»; «Мы нынче растеряли васильки. Да, сорняки... Но – до чего красивы!»).
Прикладной (ресурсный) интерес к проблеме биоразнообразия связан с использованием живых организмов для производства продуктов питания, лекарств, одежды, строительных материалов и пр. При этом из установленных 250 тыс. видов цветковых растений 3 тыс. имеют пищевое значение, около 200 – освоены и только 20 из них имеют наибольшее экономическое значение.
Экономический (и, соответственно, политический) интерес к проблеме биоразнообразия также вполне понятен.
Во-первых, биоразнообразие само по себе есть материальный ресурс – обитающие в дикой природе организмы могут представлять ценность для селекции и служить источниками тех или иных веществ, используемых в фармакологии, пищевой промышленности, парфюмерии и т.п. Один из наиболее известных примеров такого рода – препарат из дикого, произрастающего на о. Мадагаскар, вида барвинка (Catharanthus roseus), оказавшийся очень эффективным против детской лейкемии и принесший материальную выгоду, оцениваемую уже сотнями миллионов долларов!
Во-вторых, понятие биоразнообразия играет в некотором смысле знаковую роль, поскольку оказывается символом "наиболее разнообразного“ биома тропических лесов, которые, согласно популярному (хотя и не всегда верному) мнению, имеют ключевое значение для формирования газового режима атмосферы Земли.
Пищевые цепи и пищевая сеть. В функционирующей природной экосистеме не существует отходов. Все организмы, живые или мертвые, потенциально являются пищей для других организмов: гусеница ест листву, дрозд питается гусеницами, ястреб способен съесть дрозда. Когда растения, гусеница, дрозд и ястреб погибают, они в свою очередь перерабатываются редуцентами.
Пищевая цепь – последовательность организмов, в которой каждый из них съедает или разлагает другой. Пищевые цепи – это также движение питательных веществ от продуцентов, консументов (травоядных, плотоядных и всеядных) к редуцентам и обратно к продуцентам.
Все организмы, пользующиеся одним типом пищи, принадлежат к одному трофическому уровню (от греческого слова trophos – «питающиеся»).
Организмы природных экосистем вовлечены в сложную сеть многих связанных между собой пищевых цепей. Такая сеть называется пищевой сетью.
Движение энергии в экосистемах происходит посредством двух связанных типов пищевых сетей: пастбищной и детритной.
В пастбищной пищевой сети живые растения поедаются фитофагами, а сами фитофаги являются пищей для хищников и паразитов.
В детритной пищевой сети отходы жизнедеятельности и мертвые организмы разлагаются детритофагами и деструкторами до простых неорганических соединений, которые вновь используются растениями.
Молекулярные механизмы регуляции развития.
Билет № 2
Биотехнология как наука и отрасль современного производства.
Существуют несколько трактовок термина биотехнология. Часто под биотехнологией понимается наука, изучающая возможности использования организмов, биологических процессов и систем в производстве, включая превращение различных видов сырья в высококачественные продукты. Ее основой являются генетическая и клеточная инженерия в сочетании с микробиологическим синтезом и широким набором методов биохимии, биоорганической химии и биопроцессорной инженерии.
Кроме того, биотехнология трактуется как совокупность промышленных методов, использующих для производства живые организмы и биологические процессы, например в хлебопечении, виноделии, производстве медпрепаратов (эндокринных и т.п.), биологической очистке сточных вод и т.д.
Впервые термин "биотехнология" применил венгерский инженер Карл Эреки в 1917 г. Однако отдельные элементы биотехнологии появились достаточно давно. Фактически, это были попытки использовать в промышленном производстве отдельные клетки (микроорганизмы) и некоторые ферменты, способствующие протеканию ряда химических процессов.
Современную биотехнологическую промышленность иногда разделяют на четыре направления:
«Красная» биотехнология – производство биофармацевтических препаратов (протеинов, ферментов, антител) для человека, а также коррекция генетического кода.
«Зеленая» биотехнология – разработка и внедрение в культуру генетически модифицированных растений.
«Белая» биотехнология – производство биотоплива, ферментов и биоматериалов для различных отраслей промышленности.
«Академическая и правительственная» биотехнология – проведение НИР как стадия, предшествующая промышленному освоению, например, расшифровка генома риса.
Биологические технологии (биотехнологии) обеспечивают управляемое получение полезных продуктов для различных сфер человеческой деятельности. Эти технологии базируются на использовании каталитического потенциала различных биологических агентов и систем – микроорганизмов, вирусов, растительных и животных клеток и тканей, а также внеклеточных веществ и компонентов клеток.
Человек использовал биотехнологию многие тысячи лет: люди занимались пивоварением, пекли хлеб, получали кисломолочные продукты,применяли ферментации для получения лекарственных веществ и переработки отходов. Но только новейшие методы биотехнологии, включая методы генетической инженерии, основанные на работе с рекомбинантными ДНК, привели к «биотехнологическому буму», свидетелями которого являемся мы в настоящее время. Новейшие технологии генетической инженерии позволяют существенно усовершенствовать традиционные биотехнологические процессы, а также получать принципиально новыми, ранее недоступными способами разнообразные ценные продукты.
В современной биотехнологии как отрасли промышленности в соответствии со спецификой сфер ее применения целесообразно выделить в качестве самостоятельных ряд разделов следующие:
Промышленная микробиология;
Медицинская биотехнология;
Технологическая биоэнергетика;
Сельскохозяйственная биотехнология;
Биогидрометаллургия;
Инженерная энзимология;
Клеточная и генетическая инженерия;
Экологическая биотехнология.
Биотехнология как наука тесно стыкуется с рядом научных дисциплин, осуществляя их практическое применение или же являясь их основным инструментом.
Биотехнологические объекты находятся на разных ступенях организации:
а) субклеточные структуры (вирусы, плазмиды, ДНК митохондрий и хлоропластов, ядерная ДНК);
б) бактерии и цианобактерии;
в) грибы;
г) водоросли;
д) простейшие;
е) культуры клеток растений и животных;
ж) растения – низшие (анабена-азолла) и высшие – рясковые.
Лидерами биотехнологии являются сегодня США и Япония, накопившие многолетний опыт биотехнологий для сельского хозяйства, фармацевтической, пищевой и химической промышленности. Прочное положение в производстве ферментных препаратов, аминокислот, белка, медикаментов занимают страны Западной Европы (ФРГ, Франция, Великобритания), а также Россия. Эти страны характеризуются мощным потенциалом новой техники и технологии, интенсивными фундаментальными и прикладными исследованиями в различных областях биотехнологии.
Биотехнология в Беларуси
Проектная мощность опытно-промышленного производства в пересчете на жидкие препараты - около 30 т в год. С учетом того, что значительная часть выпущенной продукции будет использована в качестве посевного материала в промышленном производстве, фактические объемы выпуска биопрепаратов в республике составят около 1000 т.
Это позволит снизить импортозависимость по жизненно важным биотехнологическим продуктам и негативные последствия экологических проблем. Так, будет обеспечена 100% потребность республики в ферментном препарате Глюкозооксидаза, используемом в качестве основы биосенсоров Глюкосен для экспресс-анализа глюкозы в крови больных сахарным диабетом; в биоконсерванте Лаксил для силосования растительного сырья; в кисломолочной кормовой добавке ДКМ, пробиотических препаратах Билавет и Бацинил; будет расширено производство биопестицидов и биоудобрений, что заложит надежный фундамент экологизации сельского хозяйства, как необходимого условия вхождения республики в ВТО.
В Беларуси со времен Советского Союза осталась довольно развитая микробиологическая промышленность, однако многие заводы пришли в упадок за неимением спроса и нуждаются в коренной модернизации. С 2007 г. в стране принята Государственная программа «Биотехнология», одним из разделов которой является выведение биотехнологической промышленности на современный уровень, обеспечение ее разработками отечественной биотехнологической науки. Сейчас она влилась в новую ГП «Инновационные биотехнологии» на 2010-2012 гг. и на период до 2015 г.
Принципы и методы изучения генома человека. Проект «Геном человека».
Принципы и методы генетического и физического картирования генома человека.
Сегодня еще не разработана четкая классификация методов картирования. Так, например, одни авторы относят цитогенетические методы (FISH, PRINS и т.п.) к генетическим, другие – к физическим. По сути, все методы являются генетическими, так как конечный результат картирования – получение максимально подробной карты взаимного расположения структурных, функциональных и полиморфных последовательностей генома и определение расстояний между ними.
Генетическое картирование основано на методах классической генетики –определении групп сцепления, частоты рекомбинации и построении генетических карт, где единицей измерения служат проценты рекомбинации, или сантиморганы (сМ).
Цитогенетическое картирование осуществляется с применением методов цитогенетики, когда для локализации каких-либо нуклеотидных последовательностей и определения их взаимного расположения используются цитологические препараты.
Физическое картирование – это обширная группа молекулярно-биологических методов, позволяющая строить карты генома (обычно их называют физическими) высокого уровня разрешения и определять расстояния между локализуемыми нуклеотидными последовательностями с точностью от нескольких десятков тысяч п.н. до одной нуклеотидной пары.
Генетическое картирование. До недавнего времени изучение геномов как человека, так и других млекопитающих, было возможно только путем генетического анализа – построения генетических карт или карт сцепления (linkage map). Генетической картой хромосомы называют относительное положение генов, находящихся в одной группе сцепления. Сначала формируются группы сцепления генов и исследуется их взаимное расположение. Основным методом построения карт сцепления является классический генетический анализ, т.е. анализ наследования признаков в родословной, а также изучение частоты рекомбинации генных локусов в мейозе. Карты сцепления показывают порядок линейного расположения генов и маркеров на хромосоме и генетическое расстояние между ними, выраженное в процентах рекомбинации – сантиморганах (сМ). Считается, что два гена на хромосоме находятся на расстоянии 1 сМ, если вероятность рекомбинации между ними в процессе мейоза составляет 1%. Карта генетического сцепления составляет около 2809 сМ для мужчин и 4782 сМ для женщин. Меньший "размер" мужского генома объясняется тем, что частота рекомбинации в сперматогенезе меньше, чем в оогенезе. Средняя длина генома человека в единицах генетического расстояния составляет около 3300 сМ. Сопоставив эту величину с размером гаплоидного генома человека, оцениваемым примерно в 2,91 млрд. п.н., можно заключить, что на 1 сМ генетической карты приходится в среднем немногим менее 1 млн. п.н. ДНК на физической карте генома.
До начала 70-х гг. ХХ в. построение генетических карт человека шло очень медленными темпами. Небольшое количество детей, длительный период одного поколения, ограниченное число информативных родословных и отсутствие методов эффективного цитогенетического анализа всех пар хромосом затрудняли этот процесс. Первый ген человека был локализован на Х-хромосоме в 1911 г., а первый аутосомный ген – только в 1968 г. К середине 70-х гг. было локализовано менее 100 генов, значительная часть из них располагалась в Х-хромосоме.
Прогресс в картировании генома человека связан с мутантными генетическими линиями животных (в большинстве случаев это мыши), моделирующими различные наследственные заболевания человека.
Высокий процент сходства по нуклеотидным последовательностям между кодирующими областями гомологичных генов млекопитающих и человека, а также большое количество консервативных групп сцепления с идентичным расположением генов, так называемых синтенных групп сцепления, позволяет проводить параллельные исследования на модельных объектах, значительно ускоряющие эффективность картирования и молекулярного анализа индивидуальных генов человека.
Дальнейший прогресс в области генетического картирования связан с деятельностью крупных научно-исследовательских центров по созданию банков клеточных культур, представляющих наиболее интересные и обширные родословные. Так, в Центре по изучению полиморфизма человека – CEPH (от фр. Centre d'Etudes du Polymorphysme Humain) создана уникальная коллекция перевиваемых клеточных культур, полученных от членов семей, многоступенчатые родословные которых насчитывают десятки и даже сотни индивидуумов.
СЕРН-коллекции представляют собой идеальные системы для генетического анализа наследственных признаков. Эти коллекции были использованы исследователями во всем мире для локализации генов человека и различных типов маркеров. Были определены генотипы членов СЕРН-семей одновременно по тысячам полиморфных локусов и построены соответствующие генетические карты.
Огромный вклад в систематизацию и обобщение информации о генетических картах хромосом человека, о локализации и функциях отдельных генов и о структуре генома в целом вносят исследования, проводимые на протяжении последних 30 лет в Университете Джонса–Хопкинса в Балтиморе под руководством профессора Виктора Мак-Кьюсика. Результатом этих исследований является систематическое издание энциклопедий под названием: "Менделевское наследование у человека: Каталог аутосомно-доминантных, аутосомно рецессивных и сцепленных с Х-хромосомой фенотипов".
Цитогенетическое картирование. Методы дифференциального окрашивания позволяют идентифицировать на препарате как отдельную хромосому, так и любой участок хромосомы, выявляя окрашенные зоны – так называемые бэнды. На метафазных хромосомах малой степени спирализации идентифицируются около 750 бэндов, на прометафазных хромосомах 2500–3000. Уже разработаны методы многоцветной окраски – multicolor banding (до 25 цветов) интерфазных и метафазных хромосом.
Цитогенетические карты показывают локализацию маркера с точностью до определенной хромосомы, плеча или хромосомного сегмента. Этот тип карт обозначает линейный порядок маркеров в хромосоме. По разрешающей способности они занимают промежуточное положение между генетическими и физическими картами.
Определение хромосомной и субхромосомной локализации маркеров проводят с использованием гибридов соматических клеток между различными видами млекопитающих или непосредственной гибридизации in situ уникальных молекулярных зондов на митотические хромосомы. К методам цитогенетического картирования относятся также хромосомный сортинг (проточная цитометрия), микродиссекции и микроклонирование определенных геномных фрагментов и сравнительное генетическое картирование (сравнительная цитогенетика).
В 90-е гг. метод гибридизации in situ получил развитие в модификациях: FISH (гибридизация in situ с использованием флюоресцентной метки) и PRINS (метод, сочетающий гибридизацию in situ на метафазных хромосомах специфических праймеров с последующей ПЦР, включающей меченый биотином нуклеотид в продукт амплификации(увеличение числа копий гена в клетке)). Этот подход стал основным методом для построения цитогенетических карт.
Физическое картирование. В отличие от генетических карт, построенных на основе групп сцепления и дающих статистические расстояния между ДНК-маркерами и генами, физическое картирование позволяет определять физические расстояния между маркерами в каждой хромосоме.
К методам физического картирования относят
-рестрикционное картирование,
-RH-картирование, клонирование в YAC (от англ. yeast artificial chromosome),
-BAC (от англ. bacterial artificial chromosome),
-на космидах (гибридные векторы, как плазмиды, но обладают способностью упаковываться in vitro в капсиды фага λ), плазмидах (кольцевая внехромосомная ДНК, способная к автономной репликации) и других векторах и
-контиг-картирование на их основе, а также
-секвенирование ДНК.
Использование искусственных хромосом создает основу для проведения физического картирования как на хромосомном, так и на субхромосомном уровне.
Основой физического картирования генома является построение физических карт, т.е. определение порядка расположения физических маркеров вдоль молекулы ДНК. В качестве физических маркеров могут выступать сами гены, анонимные фрагменты ДНК (D-сегменты), точки расщепления ДНК рестриктазами и т.д. Однако при развитии работ по физическому картированию исследователи столкнулись с трудностями совмещения данных.
Для преодоления этой проблемы в 1989 г. было предложено стандартизовать все обозначения меченных последовательностей ДНК в геноме, включая все типы последовательностей, будь то просто картированный сегмент ДНК с неизвестной функцией (D-сегменты), последовательность с необычными сайтами рестрикции, проба, выявляющая полиморфизм, последовательность, гибридизующаяся с определенным "бэндом" при гибридизации in situ или STS-маркеры (от англ. sequenсed tagged site).
Контиг-карты хромосом человека на основе перекрыващихся клонов геномной ДНК человека относятся к методам высокого разрешения. Контиговая карта, или контиг (от англ. contigous – протяженный), включает набор перекрывающихся клонированных фрагментов геномной ДНК в комплексе с информацией о порядке их перекрывания. Локализовать клонированный фрагмент можно, применяя методы гибридизации с пробами ДНК (локализация которых известна), ПЦР-анализа, секвенирования.
Картирование методом дробовика (ShotGun) и картирование с использованием случайных STS предполагает секвенирование коротких случайных фрагментов генома человека и подбор ПЦР-праймеров к этим фрагментам, т.е. создание STS. Используя радиационные гибриды или иные системы, в том числе соматические гибриды и семьи полиморфных маркеров CEPH, можно определить положение STS на физической и генетической картах генома человека. Недостаток данного подхода – невозможность оценить истинное расстояние между STS-маркерами, что сильно снижает разрешающую способность получаемых карт.
Картирование с использованием панелей радиационных гибридов соматических клеток (RH-картирование, radiation hybrids mapping) сегодня получило очень широкое распространение и применяется исследователями в различных целях: для тотального картирования, для составления карт индивидуальных хромосом или участков хромосом человека на основе хромосом-специфичных RH-панелей, для установления мест локализации конкретных генов или STS, EST и других типов маркеров.
Проект «Геном человека».
В 1988 г. один из первооткрывателей двойной спирали ДНК, нобелевский лауреат Джеймс Уотсон, публично высказал мысль о том, что наука вплотную приблизилась к раскрытию химической основы наследственности "царя природы" – человека. К тому времени было уже известно, что наследственный аппарат человека, геном, т.е. совокупность всех генов и межгенных участков ДНК, составляет около 3 млрд. пар нуклеотидов. Эта величина казалась столь большой, что сама мысль о возможности получения такого объема информации представлялась совершенно фантастической.
Противники расшифровки генома считали поставленную задачу нереальной, поскольку ДНК человека в десятки тысяч раз длиннее молекул ДНК вирусов или плазмид. Главный аргумент противников сводился к тому, что «проект потребует миллиарды долларов, которых недосчитаются другие области науки; поэтому геномный проект затормозит развитие науки в целом. А если все-таки деньги найдутся и геном человека будет расшифрован, то полученная в результате информация не оправдает затрат...» Однако Дж. Уотсон остроумно парировал: «лучше не поймать большую рыбу, чем не поймать маленькую». Аргумент был услышан – проблему генома вынесли на обсуждение в конгресс США, и в итоге была принята национальная программа «Геном человека».
Одновременно с Дж. Уотсоном (в том же 1988 г.) с аналогичной идеей выступил выдающийся российский молекулярный биолог и биохимик, академик А.А.Баев (1904–1994). После консультаций с коллегами он обратился к М.С. Горбачеву с письмом, в котором предложил организовать государственный научный проект по изучению генома человека. В России, как и за ее пределами, эта идея также была встречена весьма критически, однако время шло, и очень скоро научное сообщество во всем мире стало обсуждать ее всерьез. С 1989 г. и в США, и в СССР функционируют соответствующие научные программы; позднее возникла Международная организация по изучению генома человека (HUGO), вице-президентом которой несколько лет был академик А.Д. Мирзабеков.
В результате 26 июня 2000 г. на совместной пресс-конференции с участием президента США и премьер-министра Великобритании представители двух исследовательских групп – International Human Genome Sequencing Consortium (IHGSC) и «Celera Genomics» – объявили о том, что работы по расшифровке генома человека успешно завершены, и черновой его вариант составлен. Начался новый эпизод развития человечества – постгеномная эра.
Создание первого «чернового» варианта генома человека обошлось в 300 млн. долл. Однако на все исследования по этой теме, включая сравнительные анализы и решение ряда этических проблем, было израсходавано около 3 млрд. долл. «Celera Genomics» вложила примерно столько же, правда, она истратила их всего за шесть лет. Цена колоссальная, но эта сумма ничтожна по сравнению с выгодой, которую получит страна-разработчик от ожидаемой вскоре окончательной победы над десятками серьезных заболеваний. А уже в 2007 г. Дж. Уотсону были подарены два DVD-диска с его геномом общей стоимостью в 1 млн. долл.
Российский проект «Геном человека» (РПГЧ). Одновременно с США в СССР по решению правительства было открыто финансирование и организован Научный совет по программе "Геном человека" под руководством А.А. Баева. Расположившийся в головном учреждении программы – Институте молекулярной биологии им. В.А. Энгельгардта РАН, совет весьма быстро создал инфраструктуру, объединил исследования многих разрозненных групп, преодолев ведомственные барьеры и географическую удаленность. В настоящее время в программу вовлечены 90 независимых исследовательских групп из 30 различных научных организаций. Основными целями программы являются физическое и функциональное картирование генома человека, разработка молекулярной диагностики наследственных и злокачественных заболеваний, компьютерный анализ генома (биоинформатика) поиск новых генов человека и выявление их функции.
Вопреки ограниченной финансовой поддержке Российский проект «Геном человека» успешно выполняется благодаря следующим факторам:
широкое и постоянное международное сотрудничество с ведущими лабораториями Европы и США, тесные и эффективные контакты с HUGO;
четкая организация работы Научным советом; кооперация и координация деятельности исследовательских групп в России, открытая конкуренция за гранты между исследовательскими группами, регулярное и быстрое распространение информации через электронные средства коммуникации;
в РПГЧ работают многие одаренные компьютерщики; созданы два информационных центра (проф. Ю.П.Лысова, Москва и проф. Н.А.Колчанова, Новосибирск).
Особое внимание РПГЧ уделяет медико-генетическим аспектам изучения генома. Разработана и введена в медицинскую практику в ряде регионов России молекулярно-генетическая диагностика отдельных широко распространенных заболеваний.
Секвенирование в проекте «Геном человека». Важная составляющая проекта «Геном человека» – разработка множества новых, революционизирующих методов исследований (таблица 1). Развитые еще до начала выполнения проекта методы (их назвали методами первого поколения) включали применение рестрикционных ферментов; создание гибридных молекул, их клонирование и перенос участков ДНК с помощью векторов в клетки-доноры (чаще всего в клетки кишечной палочки Escherichia coli и дрожжевые клетки; иногда этот процесс называют амплификацией генов in vivo); синтез ДНК на матрицах информационной РНК (сДНК-синтез); методы секвенирования генов; получение практически неограниченного количества копий генов с помощью PCR-машин (амплификация участков ДНК in vitro); методы, предназначенные для разделения молекул ДНК по плотности, массе, различной вторичной структуре.
Таблица 1 – Автоматизированные методы анализа последовательностей ДНК (методы секвенсового анализа), разработанные в рамках проекта «Геном человека»
Технологии секвенирования второго поколения |
Технологии секвенирования третьего поколения |
Высоковольтный капиллярный электрофорез |
Ускоренное флуоресцентное детектирование отдельных меченых оснований в проточных цитофотометрах |
Ультратонкий высоковольтный электрофорез |
Прямое чтение оснований в последовательностях ДНК с использованием сканирующих туннельных микроскопов или микроскопов, работающих на уровне атомного и субатомного разрешений |
Резонансная ионизационная спектроскопия, предназначенная для детектирования меток, созданных с помощью стабильных изотопов |
Улучшенные методы прямого масс-спектрометрического анализа последовательностей ДНК Секвенирование путем гибридизации изучаемых высокомолекулярных отрезков ДНК с панелями (ДНК-чипами) коротких отрезков нуклеотидов |
Билет № 3
Свободнорадикальные реакции в клетках и проблемы их регуляции.
Что такое свободные радикалы?
Хорошо известно, что в органических молекулах (включая те, из которых состоит наш организм) электроны на внешней электронной оболочке располагаются парами: одна пара на каждой орбитали
Свободные радикалы отличаются от обычных молекул тем, что у них на внешней электронной оболочке имеется неспаренный (одиночный) электрон
Неспаренный электрон в радикалах принято обозначать точкой. Например, радикал гидроксила обозначают как HO·, радикал перекиси водорода как HOO·, радикал супероксида как ·OO- или O2·-.
РАДИКАЛЫ-ЭТО МОЛЕКУЛЫ С НЕСПАРЕННЫМИ ЭЛЕКТРОНАМИ
Супероксид
Молекулярный кислород служит конечным акцептором электронов в митохондриальной дыхательной цепи
ЕГО восстановление описывается уравнением
O2+4e- + 4H+ 2H2O
Однако восстановление кислорода может привести к образованию не только безопасной воды
Cуществует вероятность одноэлектронного восстановления
![]()
В этом случае неспаренный электрон может атаковать ковалентную связь
Супероксид может образовываться по нескольким причинам
(1) Утечка электронов с промежуточных переносчиков на кислород при нормальном процессе восстановления молекулярного кислорода в дыхательной цепи
(2) Мутации митохондриальной ДНК могут сопровождаться блокированием электрон-транспортных путей в результате чего электроны будут расходоваться на образование супероксида (такого рода мутации особенно опасны из-за отсутствия у митохондриальных ДНК систем репарации
О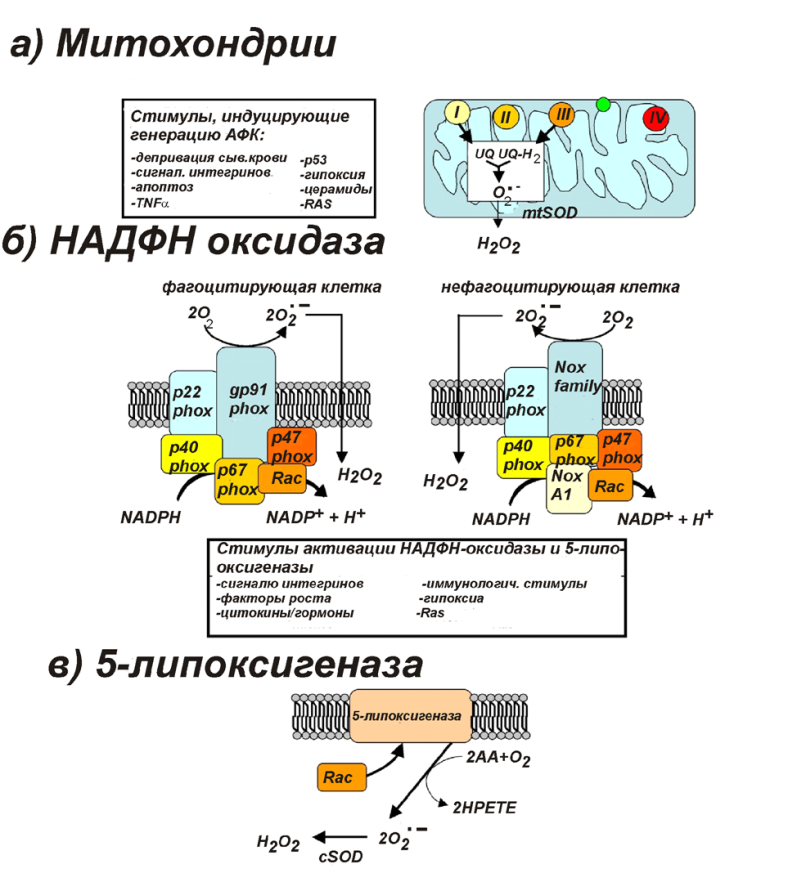 бразование
АФК
бразование
АФК
АФК (РФК) постоянно образуются в живой клетке как продукты нормального метаболизма кислорода. Активные формы кислорода образуются также под действием ионизирующего излучения. Некоторые АФК могут играть роль медиаторов важных внутриклеточных сигнальных путей. Однако, повышенная продукция АФК приводит к окислительному (оксидативному) стрессу. Нормальные функции АФК включают индукцию иммунной системы и мобилизацию систем ионного транспорта. Например, клетки крови на месте повреждения начинают продуцировать АФК, что рекрутирует тромбоциты, необходимые для начала процесса заживления раны. АФК также запускают программируемую клеточную смерть (апоптоз).
Около 95% от всего потребляемого кислорода клетки восстанавливается в митохондриях до воды в процессе окислительного фосфорилирования. Остальные 5% кислорода в результате различных реакций (как правило ферментативных) превращаются в АФК.
Супероксид сам по себе не очень токсичен и опасен, но может превращаться в опасные формы. pKa протонированного супероксида 4,8, поэтому при физиологических pH он существует как H2O2
Если повреждение значительное клетка подвергается апоптозу
![]()
(4) Ионизирующая радиация
(5) Фагоциты, захватывая бактериальные клетки потребляют много O2. Он восстанавливается с помощью NADPH с образованием .O2-
При этом он попадает в вакуоль, превращается в H2O2, которая разрушает бактериальные клетки
Количество супероксида очень мало НО !!!
Поскольку он радикал, то превращается только в радикал
В результате –цепная реакция, которая может прекратиться либо вследствие рекомбинации с другими радикалами, либо из-за низкой реакционной способности дочерних радикалов. Полагают, что это может приводить к старению, к образованию катаракты хрусталика, инфаркту миокарда.
Три способа защиты от пероксидов
Один химический и два ферментативных.
Химический способ защиты
Антиоксиданты
Антиоксиданты-вещества, которые вступая в реакцию со свободными радикалами, превращаются в вещества с малой реакционной способностью.
В результате прерывается цепной процесс
Природные тушители цепных реакций
Аскорбиновая кислота (витамин С), (растворима в воде), -токоферол (витамин Е), (растворим в жирах).
Поэтому в тандеме способны защитить и компоненты цитозоля и мембранные липиды, -каротин, Мочевая кислота
Билирубин - продукт расщепления гема гемоксигеназой. (активируется продуктами частичного восстановления кислорода (их накопление называют окислительным стрессом, может быть вызван ионизирующим излучением) и поэтому иногда производство билирубина рассматривают как ответ на окислительный стресс.
Ферментативные способы защиты Супероксиддисмутаза (СОД)
СОД катализирует реакцию
2.О2-+2H+ H2O2+O2
Разложение H2O2 катализирует каталаза
2H2O2 2H2O+ О2
Перекись водорода появляется не только в результате действия СОД
H2O2
Но также в результате действия FAD (флавинадениндинуклеотид)-содержащих оксидаз, которые катализируют реакции типа
AH2+O2 A+H2O2
Перекись водорода опасна тем, что в присутствии ионов тяжелых металлов(например, Fe2+) она распадается с образованием агрессивных гидроксильных радикалов (не путать с безобидными гидроксильными анионами
H2O2+Fe2+ Fe3++.OH+OH-
Помимо тандема супероксиддисмутазы и каталазы, разрушающих радикалы, существует еще один мощный фермент, спасающий от H2O2
ГЛУТАТИОНПЕРРОКСИДАЗА
(Защита важна для мозга где мало каталазы)
Глутатион (тиол-содержащий трипептид) -глутамил-цистеинил-глицин GSH (восстановленный глутатион)
Роль глутатиона в клетке
Глутатион содержит необычную пептидную связь между амино-группой цистеина и карбокси-группой боковой цепи глутамата. Важность глутатиона в клетке определяется его антиоксидантными свойствами. Фактически глутатион не только защищает клетку от таких токсичных агентов как свободные радикалы, но и в целом определяет редокс-статус внутриклеточной среды.
В клетке тиоловые группы находятся в восстановленном состоянии (SH) в концентрации около 5 мМ. Фактически, такая высокая концентрация глутатиона в клетке приводит к тому, что он восстанавливает любую дисульфидную связь (S-S), образующуюся между цистеинами цитозольных белков. При этом восстановленная форма глутатиона GSH превращается в окисленную GSSG. Восстанавливается окисленный глутатион под действием фермента глутатионредуктаза, которая постоянно находится в клетке в активном состоянии и индуцируется при оксидативном стрессе. Отношение восстановленный/окисленный глутатион внутри клетки является одним из важнейших параметров, который показывает уровень внутриклеточной токсичности (уровень оксидативного стресса).
Соотношение окисленного и восстановленного глутатиона в клетке
В здоровой клетке восстановленный глутатион 90%
Окисленный глутатион 10%
Функции глутатиона. Глутатион важен в качестве гидрофильной молекулы, которая присоединяется ферментами печени к гидрофобным токсическим веществам в процессе их биотрансформации с целью выведения из организма (в составе желчи). Как часть глиоксалазной ферментативной системы глутатион участвует в реакции детоксификации метилглиоксаля, токсического побочного продукта метаболизма. Глиоксалаза I превращает метилглиоксаль и восстановленный глутатион в лактоилглутатион. Глиоксалаза II гидролизует лактоилглутатион на глутатион и лактат (молочную кислоту)
Глутатион является субстратом реакций конъюгирования и восстановления, катализируемых глутатион-S-трансферазой в цитозоле, микросомах и в митохондриях.
Глутатион плохо всасывается желудочно-кишечным трактом, поэтому для восстановления нормального уровня глутатиона в случае патологии назначают N-ацетилцистеин, участвующий в его биосинтезе
ГЕНЕРАЦИЯ АКТИВНЫХ ФОРМ КИСЛОРОДА В КЛЕТКЕ
НАДФ•H-оксидаза, или NADPH-оксидаза (NOX) — клеточный мембрано-связанный мультимолекулярный ферментный комплекс, локализующийся на плазматической мембране и в некоторых органеллах. Особенно обогащены этим ферментом фагоцитарные клетки, такие как макрофаги. Эти оксидазы участвуют в клеточной противомикробной защитной системе, а также в клеточной пролиферации, дифференцировке и регуляции экспрессии генов. Существует целая группа NADPH-оксидаз, которые различаются по составу субъедениц, клеточной специфичности, регуляции и другим параметрам.
Биотестирование как составной элемент в системе оценки состояния окружающей среды.
Основные методы контроля химического загрязнения биосферы.
Контроль за загрязнением осуществляется двумя способами: посредством контроля на входе и контроля на выходе.
• Контроль на входе препятствует проникновению потенциального загрязнителя в окружающую среду или резко сокращает его поступление. Например, примеси серы могут быть удалены из угля до его сжигания. Это предотвратит или резко снизит выбросы такого загрязнителя атмосферы, как диоксид серы, химического вещества, вредного для растений и нашей дыхательной системы.
• Контроль на выходе направлен на ликвидацию отходов, уже попавших в окружающую среду. Для оценки степени загрязнения используют понятие ПДК – предельно допустимой концентрации вещества (концентрация выше ПДК – опасно, ниже – не опасно), но этого не достаточно, т.к. не учитывается воздействие нескольких веществ (синергический эффект).
Известно, что менее 10% добываемого сырья превращается в готовую продукцию; остальное - отходы, загрязняющие природную среду. Уже сегодня в банке данных Chemical Abstract Services (США) имеются сведения о почти 8 млн. различных химических соединений, причем несколько десятков тысяч из этого количества находят широкое применение в многообразных сферах жизни и постоянно используются людьми.
Основные методы контроля химического загрязнения биосферы
Экологически опасные факторы (ЭОФ химической природы – это химические вещества, которые в определенных концентрациях приводят к качественным и количественным нарушениям в экосистемах. К ЭОФ химической природы относятся, в первую очередь, следующие группы веществ:
- тяжелые металлы,
- диоксины и диоксиноподобные соединения,
- ДДТ и другие пестициды,
- нитриты, нитраты и нитрозосоединения,
- полициклические ароматические углеводороды.
Экологически опасные факторы (ЭОФ химической природы – это химические вещества, которые в определенных концентрациях приводят к качественным и количественным нарушениям в экосистемах. К ЭОФ химической природы относятся, в первую очередь, следующие группы веществ:
- тяжелые металлы,
- диоксины и диоксиноподобные соединения,
- ДДТ и другие пестициды,
- нитриты, нитраты и нитрозосоединения,
- полициклические ароматические углеводороды.
Необходим качественно новый подход к описанию состояния окружающей среды как динамической химико-биологической системы. Необходимы данные о том, как ведут себя, какие испытывают превращения, к каким последствиям приводят те или иные химические вещества, попадающие в биосферу. При этом традиционные методы химического и биологического анализов служат основой для количественной оценки состояния и динамических характеристик природных экосистем.
Изменения в природе контролируют и изучают многочисленные службы: гидрометеорологическая, сейсмическая, ионосферная, магнитометрическая и др. Но они не ставят своей основной целью выделение антропогенных сдвигов в природе на фоне естественных изменений. Поэтому и появилась потребность в специальных наблюдениях за изменениями в окружающей среде, вызванных деятельностью человека.
Мониторинг – наблюдение, оценка и прогноз состояния окружающей среды в связи с хозяйственной деятельностью человека. В системе мониторинга различают три уровня: санитарно-токсикологический, экологический и биосферный мониторинг. В настоящее время более или менее развита система санитарно-токсикологического мониторинга.
Санитарно-токсикологический (химический) мониторинг – наблюдение за состоянием окружающей среды, степенью загрязнения природных объектов вредными веществами, за влиянием этих загрязнений на человека, животный и растительный мир, за наличием в окружающей среде аллергенов, патогенных микроорганизмов, пыли т.д., за содержанием в атмосфере оксидов азота и серы, СО, тяжелых металлов, за качеством водных объектов, степенью их загрязнения органическими веществами, нефтепродуктами и минеральными солями.
Экологический мониторинг – наблюдение за изменениями в экологических системах (биогеоценозы), природных комплексах, за их продуктивностью, а также динамикой запасов полезных ископаемых, водных, земельных и растительных ресурсов. Задача экологического мониторинга – обнаружение в экосистемах изменений антропогенного характера (на фоне естественных флуктуаций).
Биосферный мониторинг – наблюдения за глобально-фоновыми изменениями в природе: степенью радиации; наличием в атмосфере СО2, О3; ее запыленностью; циркуляцией тепла; газовым обменом между океаном и воздушной оболочкой земли; мировой миграцией птиц, животных, растений и насекомых; погодно-климатическими изменениями на планете.
В основе мониторинга лежит система определения концентраций загрязняющих веществ в объектах окружающей среды - система эколого-аналитического контроля (ЭАК).
В сферу эколого-аналитического контроля входят следующие контролируемые объекты:
- воды - пресные, поверхностные, морские, подземные, атмосферные осадки, талые, сточные;
- воздух – атмосферный, природных заповедников (фон), городов и промышленных зон, рабочей зоны;
- почвы (загрязнения);
- донные отложения (загрязнения);
- растения, пища и корма, животные ткани (загрязнения).
В сферу объектов ЭАК при необходимости могут быть включены и другие объекты, представляющие по той или иной причине опасность для окружающей среды, в частности, полупродукты и готовая продукция нефтехимической, химической, фармацевтической и микробиологической промышленности.
Эффективность и достоверность методик и методического обеспечения системы ЭАК определяются прежде всего пробоотбором и пробоподготовкой. Технические средства, используемые при отборе проб, пробоподготовке и определении в полевых условиях, должны функционировать автономно в широком диапазоне температур (от -50 до +50) и влажности (до 100%), выдерживая агрессивные воздействия сред, в которых и с которыми они работают. В этой связи весьма перспективны химические и физические сенсоры – эффективные средства оперативного контроля газовых и жидких сред на содержание нормируемых микрокомпонентов, в которых пробоотбор и несложная пробоподготовка совмещены с собственно определением. Они особенно удобны для оснащения постов эколого-аналитического контроля, а также создания поли-функциональных автоматизированных систем контроля состояния воздушного бассейна, воздушных выбросов предприятий, природных и сточных вод.
Для решения ЭАК используют инструментальные методы современной аналитической химии, основанные на измерении различных физических свойств определяемых веществ или продуктов их химических превращений (аналитических реакций) с помощью физических и физико-химических приборов. Результат измерения, несущий химико-аналитическую информацию, часто называют аналитическим сигналом.
Ценную информацию в анализе вод предоставляют электрохимические методы анализа: потенциометрия, поляро-графические и кулонометри-ческие методы. В качестве примера приведена популярная диаграмма Eh-рН (окислительно-восстановительный по-тенциал) как результат потенцио-метрических измерений. Как видно из рисунка, эти параметры для различных вод неодинаковы. Eh и рН контролиру-ются такими важнейшими факторами, как фотосинтез дыхания и разложения, окислительно-восстановительные реак-ции с участием серы, железа и углерода, рассмотренный ранее баланс между растворенной углекислотой и карбо-натом кальция в природных водах и другие факторы химической экологии гидросферы.
Исключительно мощное средство контроля загрязнения различных объектов окружающей среды - хроматографические методы, позволяющие анализировать сложные смеси компонентов. Наибольшее значение приобрели тонкослойная, газожидкостная и высокоэффективная жидкостная и ионная хроматография. Будучи несложной по технике выполнения, тонкослойная хроматография хороша при определении пестицидов и других органических соединений-загрязнителей. Газожидкостная хроматография эффективна при анализе многокомпонентных смесей летучих органических веществ. Применение различных детекторов, например малоизбирательного детектора по теплопроводности - катарометра и избирательных - пламенно-ионизационного, электронного захвата, атомно-эмиссионного, позволяет достигать высокой чувствительности при определении высокотоксичных соединений.
Высокоэффективную жидкостную хроматографию применяют при анализе смесей многих загрязняющих веществ, прежде всего нелетучих. Используя высокочувствительные детекторы: спектрофотометрические, флуориметрические, электрохимические, можно определять очень малые количества веществ. При анализе смесей сложного состава особенно эффективно сочетание хроматографии с инфракрасной спектрометрией и особенно с масс-спектрометрией. В последнем случае роль детектора играет подключенный к хроматографу масс-спектрометр. Обычно приборы такого типа оснащены мощным компьютером. Так определяют пестициды, полихлорированные бифенилы, диоксины, нитрозоамины и другие токсичные вещества. Ионная хроматография удобна при анализе катионного и анионного составов вод.
Приборы ЭАК входят в общую совокупность аналитических приборов. Все приборы ЭАК – выпускаемые или разрабатываемые - можно разделить на две группы: приборы общего назначения и специализированные приборы.
В первую группу входят приборы, применение которых не жестко связано со спецификой контролируемого объекта или определяемого показателя, т.е. возможно их использование для большого числа методик анализа, охватывающих контроль разнотипных объектов и разнообразных показателей. К таким приборам относится большинство хроматографов, спектрофотометров, спектрографов, спектрофлуориметров, полярографов и масс-спектрометров.
Вторая группа включает приборы, предназначенные для определения конкретного компонента в конкретном объекте контроля (пример – анализатор SO2 в промышленных выбросах или анализатор нефтепродуктов в водах).
Недостатки ис¬пользования химических и других методов при оценке биобезопасности химических соединений среды в целом.
Индустриализация и урбанизация, особенно технологический прогресс и развитие агрохимического комплекса, и, как следствие, объективный процесс “химизации” среды обитания человека инициировали повышение значимости эколого-аналитического контроля. Расширились не только номенклатуры объектов и круг определяемых в них компонентов. Резко увеличилось число микро- и макрокомпонентов, одновременно присутствующих в анализируемых пробах. В такой ситуации узкопрофильные методы и приборы, предназначенные для качественного и количественного анализа только отдельных конкретных веществ, часто не обеспечивают требуемое качество результатов химического анализа вслествие их неселективности. Поэтому на передний план стали выходить многоцелевые методы и аналитические комплексы, позволяющие достоверно определять контролируемые вещества в многокомпонентных пробах.
Современным аналитическим задачам присуща ярко выраженная тенденция к снижению требуемых пределов обнаружения. Такая тенденция проявляется в эколого-аналитическом контроле содержания супертоксикантов, таких как 2,3-7,8-диоксин, 3,4-бензпирен, нитрозоамина, некоторых видов пестицидов и др. В качестве обязательного элемента эколого-аналитического комплекса, помимо пробоотборных устройств и развитой системы пробоподготовки, выступает система предварительного концентрирования определяемых компонентов, позволяющая выделять микрокомпоненты из образцов разнообразного состава.
В России установлены ПДК примерно для 1400 соединений в воде, более 1300 соединений в воздухе и свыше 200 – в почвах, т.е. только для незначительной части соединений, поступающих в окружающую среду. Следует отметить один существенный недостаток принятого подхода к эколого-аналитическому контролю, который является общим для России и других стран. Такой подход состоит в установлении ПДК соединений, разработке методик, обеспечивающих определение этих соединений в объектах окружающей среды на уровне ПДК и ниже, и их утверждение в соответствующих ведомствах. Очевидны недостатки существующего подхода: колоссальные затраты времени и высококвалифицированного труда и, как следствие, огромные экономические потери. К тому же такой подход не охватывает контроль содержания неизвестных, ненормируемых веществ и их влияния на экологическую обстановку.
Характеристика понятий биоиндикация и биотестирование.
Биомониторинг является составной частью экологического мониторинга и включает в себя такие подсистемы, как биотестирование, биоиндикацию и биоаккумуляцию. Биотестирование и биоиндикация – это два схожих исследовательских приема, в которых о качестве среды, о факторах, воздействующих на эту среду, судят по выживаемости, продуктивности, поведению, а также по различным физико-биологическим параметрам живых организмов. Различие состоит в том, что биотестирование подразумевает использование живых организмов, специально помещаемых в данную среду (тест-объекты), а биоиндикация исследует живые организмы, естественным образом обитающие в данной исследуемой среде. Биоаккумуляция – частный случай биотестирования или биоиндикации, когда о качестве среды и факторах, воздействующих на эту среду, судят по степени накопления вредных веществ в живых организмах.
Биомониторинг проводится на организменном, популяционном, экосистемном и биосферном уровнях. На организменном уровне определяется содержание различных загрязняющих веществ в живых организмах. На популяционном уровне контролируется изменение популяционных характеристик под воздействием тех или иных факторов во времени, например численности популяций, их возрастной и половой структуры и.п. На экосистемном уровне мониторинг сводится к постоянной регистрации видового состава, структурных и функциональных характеристик биологических систем, изменения площади, занимаемой ими и т.д.
Приемы и подходы при выборе тест-объектов и тест-реакций.
Биомониторинг проводится на организменном, популяционном, экосистемном и биосферном уровнях. На организменном уровне определяется содержание различных загрязняющих веществ в живых организмах. На популяционном уровне контролируется изменение популяционных характеристик под воздействием тех или иных факторов во времени, например численности популяций, их возрастной и половой структуры и.п. На экосистемном уровне мониторинг сводится к постоянной регистрации видового состава, структурных и функциональных характеристик биологических систем, изменения площади, занимаемой ими и т.д.
Можно привести ряд примеров использования методов биотестирования, Так, в Голландии в качестве тест-объектов на больших площадях страны используются различные полезные для человека растения: гладиолусы и тюльпаны являются тест-объектами на накопление фторидов; итальянская ржаная трава – тест-объект на накопление ионов тяжелых металлов. В последнее время широкое применение во многих странах находит биотестирование с помощью таких высокочувствительных к загрязнению разного рода (ионы тяжелых металлов, нефтепродукты, УФ-излучение и т.д.) тест-объектов, как дафнии (рачки – фильтраторы водоемов), пиявки, инфузории (простейшие), хлорелла (микроводоросль) и т.п.
В целях биоиндикации используется метод лихеноиндикации (от латинского «lichenes» – лишайники), основанный на учете количества лишайников в городских насаждениях, районах крупных предприятий и т.д. Установлена четкая связь между «полями загрязнения воздуха» в городах и встречаемостью лишайников на стволах деревьев. Весьма перспективным методом оценки качества поливной воды в садоводческих хозяйствах Подмосковья, Дальнего Востока и т.д. является так называемый «coli-тест» на наличие в воде бактерии кишечной палочки (Escherichia coli).
Кроветворная ткань является одним из основных тест-объектов биоиндикации воздействия антропогенных факторов на организм животных. Повышение над контрольным уровнем числа клеток с хромосомными аберрациями является классической цитогенетической тест-реакцией повреждения ДНК. Однако в случае пойкилотермных животных с невысоким темпом деления клеток микроядерный тест имеет больше преимуществ над метафазным и анафазным методами. Другой тест-реакцией, указывающей на угнетающие действие на кроветворную ткань и возможное мутагенное воздействие, является клеточная гибель.
Филогенетически различные виды-биоиндикаторы могут отличаться по чувствительности на уровне генетической детерминации процессов формирования, сохранения и элиминации клеток с хромосомными аберрациями – маркерами мутагенного воздействия. Амфибии и млекопитающие (мышевидные грызуны) являются классическими видами-биоиндикаторами. Моллюски, как мы могли видеть выше, представляют собой перспективный вид-биоиндикатор. В лаборатории моделирования генетических процессов по инициативе к.б.н. В.Ю.Афонина начаты работы по введению новой тест-системы на основе легочного брюхоногого моллюска большого прудовика (Lymnaea stagnalis).
Lymnaea stagnalis среди моллюсков относится к наиболее развитым и по многим параметрам близок к млекопитающим. Продолжительность жизни и скорость смены поколений у этого вида моллюсков близка к таковым у мышевидных грызунов, используемых в биомониторинге. С другой стороны, на онтогенез моллюсков как и амфибий, накладывает отпечаток сезонные факторы, что позволяет контролировать немутагенные параметры среды, влияющие на различные клеточные процессы. Связь частоты повреждений ДНК, индуцированных мутагенами, и генетической структуры популяции по различным ферментам, указывают на возможное сочетанное применение этих критериев в комплексе с цитогенетическими повреждениями. Возможность прижизненных наблюдений за различными биологическими характеристиками моллюсков, делает данный вид удобным для биоиндикационных исследований в лаборатории.
Визуальные и автоматизи¬рованные системы первичной оценки биобезопасности среды.
Важную роль в мониторинге и прогнозировании играют геоинформационные системы. Геоинформационная система (также ГИС – географическая информационная система) – система, предназначенная для сбора, хранения, анализа и графической визуализации пространственных данных и связанной с ними информации о представленных в ГИС объектах. Геоинформационнуюя систему "Республика Беларусь"
Среди белорусских экологических геоинформационных ресурсов можно назвать ГИС «Беловежская пуща». Этот проект охватывает Государственный национальный парк «Беловежская пуща», расположенный в Беларуси на границе с Польшей и занимающий площадь около 90 тыс. га. ГИС «Беловежская пуща» включает следующие основные блоки: топографическая основа, лесные ресурсы, геологические ресурсы, экологическая информация, растительность, животный мир, климат, управление, изображения, энциклопедия. Дальнейшее развитие ГИС «Беловежская пуща» предполагает создание в республике единой геоинформационной системы охраняемых и заповедных территорий, включающей в первую очередь Березинский биосферный и Припятский ландшафтно-гидрологический заповедники, Национальный парк «Браславские озера».
Среди автоматизированных систем контроля качества окружающей среды широкое распространение получили автоматизированные системы контроля загрязнения атмосферы АСКОС. В такие системы поступают информация от автоматических станций контроля загрязнения атмосферы и контроля турбулентности приземного слоя атмосферы, а также метеорологические данные из управления по гидрометеорологии. Далее с помощью соответствующего программного обеспечения осуществляется оценка современного состояния и дается краткосрочный прогноз уровня загрязнения атмосферного воздуха, что является исходными данными при решении вопроса о необходимости уменьшения выброса.
Один из современных методов автоматизированного дистан-ционного контроля качества окружающей среды – аэро-космический мониторинг. При этом достигается большая обзорность и одномоментность. В частности, спутник мгновенно может обеспечить съемку от 8 до 40 тыс. км2, а за 10 мин работы – до 1 млн. км2.
Принципы и организация системы химической безопасности.
Условиями эффективного достижения указанной цели, при которых химическая и биологическая безопасность обеспечивается на долгосрочную перспективу являются:
предупреждение возникновения источников и очагов химического и биологического поражения (заражения) путем систематического мониторинга химических и биологических опасностей, совершенствования законодательства и нормативных документов в области химической и биологической безопасности, а также контроля их исполнения;
уменьшение масштабов потенциальных очагов химического и биологического поражения и суммарных площадей зон защитных мероприятий путем проведения комплекса мер в отношении источников химической и биологической опасности;
повышение защищенности населения и среды его обитания от негативных влияний опасных химических веществ и биологических агентов и (или) снижение уровня их влияния путем внедрения современных средств защиты, разработанных с учетом мониторинга опасных биологических и химических факторов окружающей среды.
В Республике Беларусь необходимо разработать закон о безопасности химических веществ. По словам специалиста, в законодательной сфере есть определенные недочеты, касающиеся управления обращением химических веществ. Это связано, в первую очередь, с комплексным подходом к регулированию контроля за полным жизненным циклом химвещества. Существенным моментом, считает замдиректора РНПЦ, является классификация и маркировка химических веществ. По ее мнению, в законодательстве должно быть оговорено, каким образом проводится эта процедура, как подается информация об опасности, которую представляет химическое вещество. Она также отметила, что на сегодняшний день конкретно не обозначено, какие вещества полностью запрещены к обороту на территории республики. В стране отсутствует система контроля за распространением особо опасных токсикантов, в частности стойких органических загрязнителей. Не определена значимость уровней аварийных загрязнений в зависимости от длительности пребывания человека в зоне аварии, нет единых подходов к оценке потенциальной аварийной опасности промышленных химических объектов. В стране отсутствует система контроля за распространением особо опасных токсикантов, в частности стойких органических загрязнителей.
Индустриально-городские экосистемы и их структура.
Возникновение индустриально – городских экосистем было вызвано в основном процессами урбанизации. В современной экологии сформировались два тесно связанных между собой научных направления – экология городов (урбоэкология) и экология городского населения.
Урбанизация – это рост и развитие городов, увеличение доли городского населения в стране за счет сельской местности, процесс повышения роли городов в развитии общества. Экологические проблемы городов, главным образом наиболее крупных из них, связаны с чрезмерной концентрацией на сравнительно небольших территориях населения, транспорта и промышленных предприятий, с образованием антропогенных ландшафтов, очень далеких от состояния экологического равновесия. Темпы роста населения мира в 1.5-2.0 раза ниже роста городского населения, к которому сегодня относится 40% людей планеты.
Круговорот вещества и энергии в городах значительно превосходит таковой в сельской местности. Над крупными городами атмосфера содержит в 10 раз больше аэрозолей и в 25 раз больше газов. При этом 60-70% газового загрязнения дает автомобильный транспорт. Более активная конденсация влаги приводит к увеличению осадков на 5-10%. Самоочищению атмосферы препятствует снижение на 10-20% солнечной радиации и скорости ветра.
При малой подвижности воздуха тепловые аномалии над городом охватывают слои атмосферы в 250-400 м, а контрасты температуры могут достигать 5-6°С. С ними связаны температурные инверсии, приводящие к повышенному загрязнению, туманам и смогу.
Города потребляют в 10 и более раз больше воды в расчете на 1 человека, чем сельские районы, а загрязнение водоемов достигает катастрофических размеров. Водоносные горизонты под городами сильно истощены в результате непрерывных откачек скважинами и колодцами, а кроме того загрязнены на значительную глубину.
Коренному преобразованию подвергается и почвенный покров городских территорий. На больших площадях, под магистралями и кварталами, он физически уничтожается, а в зонах рекреаций - парки, скверы, дворы - сильно уничтожается, загрязняется бытовыми отходами, вредными веществами из атмосферы, обогащается тяжелыми металлами, обнаженность почв способствует водной и ветровой эрозии.
Растительный покров городов обычно практически полностью представлен “культурными насаждениями” - парками, скверами, газонами, цветниками, аллеями. Многолетние растения в городах развиваются в условиях сильного угнетения.
Билет № 4
Структурно-функциональные особенности митохондриального генома человека и митохондриальные заболевания.
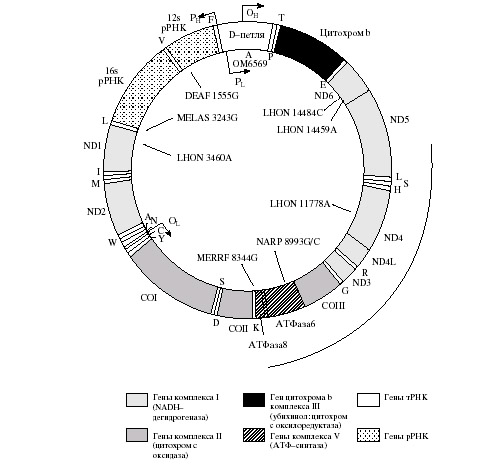
Исследование генома митохондрий выявило его совершенно уникальные особенности. Первое шокирующее сообщение пришло в 1979 г., когда было обнаружено, что в митохондриях нередки отклонения от универсального генетического кода. Эти отклонения позже были найдены во многих организмах, особенно в митохондриях животных и грибов. Так, в митохондриях человека кодон АУА кодирует аминокислоту метионин вместо изолейцина в стандартном коде. Кодоны АГА и АГГ, в стандартном коде кодирующие аргинин, являются стоп-кодонами, а кодон УГА, в стандартном коде являющийся стоп-кодоном, кодирует триптофан. Митохондрии растений, по-видимому, используют нормальный генетический код.
Митохондрии отличаются также особенностью узнавания кодонов транспортными РНК: одна молекула узнает сразу четыре кодона. Указанное изменение митохондриального генетического кода снижает значимость третьего нуклеотида в кодоне и приводит к тому, что требуется меньше транспортных РНК. Всего 22 транспортных РНК достаточно для узнавания всех 64 кодонов, тогда как для обычных рибосом их должно быть не менее 32 (в некоторых организмах найдено до 61).
Митохондриальные заболевания человека
В настоящее время известно более 100 точковых мутаций и несколько сот структурных перестроек митохондриальной ДНК (мтДНК), связанных с характерными нейромышечными и другими митохондриальными синдромами: от летальных в неонатальном периоде жизни – до заболеваний с поздним началом. Непосредственная причина возникновения и развития митохондриальных расстройств кроется в дефектах системы окислительного фосфорилирования. Благодаря выявлению все большего количества пациентов и описанию сотен родословных, сведения о взаимоотношении между генотипом и фенотипом, структуре и частоте встречаемости патогенных и условно-патогенных мутаций мтДНК в популяциях накапливаются ускоренными темпами.
Можно привести следующую классификацию патогенных мутаций мтДНК:
Смысловые мутации в структурных генах. Наиболее типичным заболеванием, ассоциированным со смысловыми заменами в митохондриальном геноме, является наследуемая по женской линии нейропатия (атрофия) зрительных нервов Лебера (LHON).
Мутации мтДНК, приводящие к нарушениям синтеза белка. Прогрессирующая с возрастом нейросенсорная глухота, если она наследуется по линии матери, часто ассоциируется с заменой А на Г в позиции 1555 гена 12S рРНК.
Структурные перестройки мтДНК. Описано множество перестроек в мтДНК (дупликации, делеции или их комбинации), приводящих к нарушениям в системе окислительного фосфорилирования. Такие перестройки обнаружены у больных нейромышечными заболеваниями типа синдрома Кернса–Сейра (KSS), при прогрессирующей наружной офтальмоплегии (СРЕО) и синдроме Пирсона (болезнь костного мозга и поджелудочной железы).
В
 заимодействие
двух геномов.
Хотя
большинство генов, продукты которых
ответственны за нормальное функционирование
системы окислительного фосфорилирования,
располагаются в хромосомах, сегодня
известно всего несколько локусов,
мутации в которых могут рассматриваться
как причина митохондриальной болезни.
В качестве примера можно сослаться на
синдром нейрогастроинтестинальной
энцефалопатии (MNGIE), обусловленной
мутацией гена тимодинфосфорилазы.
Кроме того, в основе возникновения
некоторых митохондриальных миопатий,
таких как офтальмоплегия и птоз, может
находиться дестабилизация молекулы
мтДНК, инициируемая мутациями в ядерных
генах.
заимодействие
двух геномов.
Хотя
большинство генов, продукты которых
ответственны за нормальное функционирование
системы окислительного фосфорилирования,
располагаются в хромосомах, сегодня
известно всего несколько локусов,
мутации в которых могут рассматриваться
как причина митохондриальной болезни.
В качестве примера можно сослаться на
синдром нейрогастроинтестинальной
энцефалопатии (MNGIE), обусловленной
мутацией гена тимодинфосфорилазы.
Кроме того, в основе возникновения
некоторых митохондриальных миопатий,
таких как офтальмоплегия и птоз, может
находиться дестабилизация молекулы
мтДНК, инициируемая мутациями в ядерных
генах.
Рисунок 3 – Карта мтДНК человека с указанием положения генов и относительно часто встречающихся патогенных мутаций
Изучается также генетическая предрасположенность белорусов к сердечно-сосудистым заболеваниям, ведется анализ генотипического риска в белорусской популяции по ключевым белкам липидного метаболизма и т.д. Успешно выполнен инновационный проект по геногеографии белорусов на основе анализа митохондриальной ДНК и Y-хромосомы.
Биосфера, и её основные компоненты.
Биосфера (от греч. bios - жизнь, sphaira - шар) - область системного взаимодействия живого и костного вещества планеты. Она представляет собой глобальную экосистему - совокупность всех биогеоценозов (экосистем) нашей планеты.
Та часть литосферы, гидросферы и атмосферы Земли, в которой существуют и развиваются растительные и живые организмы, называется биосферой. Иначе, биосфера- это оболочка жизни. В ее состав входят не только растительный покров и животное население планеты, все реки и озера, водная масса океанов, но и почвенный слой, значительная часть тропосферы и самый верхний слой земной коры - зоны выветривания. На земной поверхности практически нет площадей, где отсутствует жизнь. Даже в жарких и безводных тропических пустынях или на поверхности высокогорных ледников и полярных льдов обнаружены микробы и другие микроорганизмы.
Первые представления о биосфере как «области жизни» и наружной оболочке Земли были высказаны в начале 19 века Ж.Ламарком. В 1875 г. австрийский геолог Э.Зюсс впервые ввел в научную литературу современный термин «биосфера», понимая под ним область взаимодействия основных оболочек Земли: атмо-, гидро- и литосферы, где встречаются живые организмы.
Заслуга создания целостности учения о биосфере принадлежит В.И.Вернадскому. Используя этот термин, он создал науку «биосфера», ввел понятие «живое вещество» - совокупность всех живых организмов, а также отвел живым организмам роль главнейшей преобразующей силы планеты Земля, учитывая деятельность организмов не только в настоящее время, но и в прошлом. Поэтому биосфера - это все пространство, где существует или когда-либо существовала жизнь, т.е. где встречаются живые организмы или продукьы их жизнедеятельности. Ту часть биосферы, где живые организмы встречаются в настоящее время, обычно называют современной биосферой или необиосферой, а древние биосферы относят к былым биосферам, иначе палеобиосферам или мегабиосферам. Примеры последних - безжизненные ск4опления органических веществ (залежи угля, нефти, газа и др.) или запасы иных соединений, образовавшихся при непосредственном участии живых организмов (известняки, ракушечники, образования мела, ряда руд и многое другое).
Биосфера включает в себя:
• Аэробиосферу - нижнюю часть атмосферы;
• Гидробиосферу - всю гидросферу;
• Литобиосферу - верхние горизонты литосферы (твердой земной оболочки).
Границы нео - и палеобиосферы различны.
Верхняя граница теоретически определяется озоновым слоем. Для необиосферы - это нижняя граница озонового слоя (около 20 км), ослабляющего до приемлемого уровня губительное космическое ультрафиолетовое излучение, а для палеобиосферы - это верхняя граница того же слоя (около 60 км), ибо кислород в атмосфере Земли есть результат преимущественно жизнедеятельности растительности.
В большинстве случаев в качестве верхней теоретической границы биосферы указывают озоновый слой без уточнения его границ.
Практически же максимальная высота над уровнем моря, на которой может существовать жизнь, ограничены уровнем, до которого сохраняются положительные температуры и могут жить растения. Выше, до «линии снегов», обитают лишь пауки и некоторые клещи. Еще выше живые организмы могут попадаться лишь случайно.
На высотах 7500-8000м критически низкого для абсолютного большинства организмов значения достигает другой абиотический фактор - абсолютное атмосферное давление. Наиболее зависимы от величины давления птицы и летающие насекомые, преимущественно занимающие нижнюю зону.
Вся толща Мирового океана по современным представлениям полностью занята жизнью.
Нижняя граница существования активной жизни традиционно определяется дном океана 11022м (максимальная глубина Марианской впадины) и глубиной литосферы, характеризующейся температурой 100 градусов С (около 6000 м, по данным сверхглубокого бурения на Кольском полуострове). В основном жизнь в литосфере распространена лишь на несколько метров вглубь, ограничиваясь почвенным слоем.
Осадочные породы, практически все претерпевшие переработку живыми организмами, определяют нижнюю границу былых биосфер, которая, тем не менее, не опускается на материках ниже самых больших глубин океана.
Атмосфера – наиболее легкая оболочка Земли, которая граничит с космическим пространством; через атмосферу осуществляется обмен вещества и энергии с космосом.
Атмосфера имеет несколько слоев:
тропосфера – нижний слой, примыкающий к поверхности Земли (высота 9–17 км). В нем сосредоточено около 80% газового состава атмосферы и весь водяной пар;
стратосфера;
ноносфера – там “живое вещество” отсутствует.
Преобладающие элементы химического состава атмосферы: N2 (78%), O2 (21%), CO2 (0,03%).
Гидросфера – водная оболочка Земли. В следствие высокой подвижности вода проникает повсеместно в различные природные образования, даже наиболее чистые атмосферные воды содержат от 10 до 50 мгр/л растворимых веществ.
Преобладающие элементы химического состава гидросферы: Na+, Mg2+, Ca2+, Cl–, S, C. Концентрация того или иного элемента в воде еще ничего не говорит о том, насколько он важен для растительных и животных организмов, обитающих в ней. В этом отношении ведущая роль принадлежит N, P, Si, которые усваиваются живыми организмами. Главной особенностью океанической воды является то, что основные ионы характеризуются постоянным соотношением во всем объеме мирового океана.
Литосфера – внешняя твердая оболочка Земли, состоящая из осадочных и магматических пород. В настоящее время земной корой принято считать верхний слой твердого тела планеты, расположенный выше сейсмической границы Мохоровичича. Поверхностный слой литосферы, в котором осуществляется взаимодействие живой материи с минеральной (неорганической), представляет собой почву. Остатки организмов после разложения переходят в гумус (плодородную часть почвы). Составными частями почвы служат минералы, органические вещества, живые организмы, вода, газы. Преобладающие элементы химического состава литосферы: O, Si, Al, Fe, Ca, Mg, Na, K.
Ведущую роль выполняет кислород, на долю которого приходится половина массы земной коры и 92% ее объема, однако кислород прочно связан с другими элементами в главных породообразующих минералах. Т.о. в количественном отношении земная кора – это “царство” кислорода, химически связанного в ходе геологического развития земной коры.
Постепенно идея о тесной взаимосвязи между живой и неживой природой, об обратном воздействии живых организмов и их систем на окружающие их физические, химические и геологические факторы все настойчивее проникала в сознание ученых и находила реализацию в их конкретных исследованиях. Этому способствовали и перемены, произошедшие в общем подходе естествоиспытателей к изучению природы. Они все больше убеждались в том, что обособленное исследование явлений и процессов природы с позиций отдельных научных дисциплин оказывается неадекватным. Поэтому на рубеже ХIХ – ХХ вв. в науку все шире проникают идеи холистического, или целостного, подхода к изучению природы, которые в наше время сформировались в системный метод ее изучения.
Результаты такого подхода незамедлительно сказались при исследовании общих проблем воздействия биотических, или живых, факторов на абиотические, или физические, условия. Так, оказалось, например, что состав морской воды во многом определяется активностью морских организмов. Растения, живущие на песчаной почве, значительно изменяют ее структуру. Живые организмы контролируют даже состав нашей атмосферы. Число подобных примеров легко увеличить, и все они свидетельствуют о наличии обратной связи между живой и неживой природой, в результате которой живое вещество в значительной мере меняет лик нашей Земли. Таким образом, биосферу нельзя рассматривать в отрыве от неживой природы, от которой она, с одной стороны зависит, а с другой – сама воздействует на нее. Поэтому перед естествоиспытателями возникает задача – конкретно исследовать, каким образом и в какой мере живое вещество влияет на физико-химические и геологические процессы, происходящие на поверхности Земли и в земной коре. Только подобный подход может дать ясное и глубокое представление о концепции биосферы. Такую задачу как раз и поставил перед собой выдающийся российский ученый Владимир Иванович Вернадский (1863 – 1945).
Центральным в этой концепции является понятие о живом веществе, которое В.И.Вернадский определяет как совокупность живых организмов. Кроме растений и животных, В.И.Вернадский включает сюда и человечество, влияние которого на геохимические процессы отличается от воздействия остальных живых существ, во-первых, своей интенсивностью, увеличивающейся с ходом геологического времени; во-вторых, тем воздействием, какое деятельность людей оказывает на остальное живое вещество.
Билет № 5
Биотехнология в сельском хозяйстве. Землеудобрительные препараты, биопестициды, биоинсектициды, фитостимуляция. Биотехнология в животноводстве.
Биологическая защита - это использование живых организмов или продуктов их жизнедеятельности для предотвращения или уменьшения ущерба, наносимого сельскому хозяйству. В природе, в естественных экосистемах, нет ни "вредителей", ни "сорняков". Даже в агроценозах общее число полезных видов в десятки и сотни раз выше числа видов-"вредителей". Задача состоит в том, чтобы не подавлять численность естественных видов-регуляторов, а поддерживать ее, вплоть до специального насыщения агроценозов желательными видами. К достоинствам биологических методов относятся экологическая чистота, низкая энергоемкость и относительная дешевизна.
Микробные землеудобрительные биопрепараты и их использование в сельском хозяйстве
Бактериальные удобрения на основе клубеньковых бактерий, нитрагин и ризоторфин.
Микрофлора почвы оказывает непосредственное влияние на её плодородие и, как следствие, на урожайность растений. Почвенные микроорганизмы в процессе роста и развития улучшают структуру почвы, накапливают в ней питательные вещества, минерализуют различные органические соединения, превращая их в легко усвояемые растением компоненты питания. Для стимуляции этих процессов применяют различные бактериальные удобрения, обогащающие ризосферу растений полезными микроорганизмами. Микроорганизмы, используемые для производства бактериальных препаратов, способствуют снабжению растений не только элементами минерального питания, но и физиологически активными веществами (фитогормонами, витаминами и др.).
В настоящее время выпускают такие бактериальные удобрения, как нитрагин, ризоторфин, азотобактерин, фосфобактерин, экстрасол.
Российская промышленность выпускает два вида нитрагина: почвенный и сухой. Впервые культура клубеньковых бактерий на почвенном субстрате была приготовлена в 1911 году на бактериально-агрономической станции в Москве. В настоящее время его производство имеет ограниченное значение, так как технология довольно сложна и трудоёмка при выполнении отдельных операций. Более перспективна технология производства сухого нитрагина.
Препарат клубеньковых бактерий может выпускаться и в виде ризоторфина. Впервые торфяной препарат клубеньковых бактерий был приготовлен в 30-х годах, но технология была создана в 1973-77 гг.
Все виды Azotobacter строгие аэробы. Чувствительны к содержанию в среде фосфора и развиваются лишь при высоком его содержании в питательной среде. Азотфиксирующая способность культуры подавляется аммиаком (вообще содержание в среде связанного азота угнетает азотфиксацию). Стимулируют процесс фиксации азота соединения молибдена. Поэтому использовать азотобактерин рекомендуется только на почвах, содержащих фосфор и микроэлементы. Азотобактерин применяют для бактеризации семян, рассады, компостов. При этом урожайность увеличивается на 10-15%.
В нашей стране удобрение на основе азотобактера разрабатывал и исполь-зовал д.б.н. Н.А. Троицкий (лаборатория генетики микроорганизмов Института генетики и цитологии АН БССР) в конце 1980-1990-х гг.
Бактериальное удобрение фосфобактерин.
Фосфобактерин - бактериальное удобрение, содержащее споры микроорганизма Bacillus megaterium var. phosphaticum. Представляет собой порошок светло-серого или желтоватого цвета.
Бактерии обладают способностью превращать сложные фосфорорганические соединения (нуклеиновые кислоты, нуклеопротеиды и т.д.) и трудноусвояемые минеральные фосфаты в доступную для растений форму. Кроме этого бактерии вырабатывают биологически активные вещества (тиамин, пиридоксин, биотин, пантотеновую и никотиновую кислоты и др.), стимулирующие рост растения. Фосфобактерин относится к числу препаратов со стимулирующим эффектом.
Bacillus megaterium var. phosphaticum представляют собой мелкие, грамположительные аэробные спорообразующие палочки размером 2,6 мкм. Клетки содержат значительное количество соединений фосфора. В ранней стадии развития это подвижные одиночные палочки, при старении образуют эндоспоры, локализующиеся в одном из концов клетки. В силу вышеизложенного технология выращивания сводится к получению спор.
Фосфобактерин рекомендуют применять на черноземных почвах, которые содержат наиболее значительное количество фосфороорганических соединений. Необходим для повышения урожайности зерновых, картофеля, сахарной свеклы и др. сельскохозяйственных растений.
Кормовые антибиотики, антибиотики против фитопатогенов, биостимуляторы, пищевые консерванты
Антибиотики применяют в нескольких целях:
для борьбы с болезнями животных;
для борьбы с болезнями растений;
как стимуляторы роста животных;
при консервировании продуктов.
Современное определение термина "антибиотик" принадлежит М.М.Шемякину и А.С.Хохлову (1961), которые предложили считать антибиотическими веществами все продукты обмена любых организмов, способные избирательно убивать или подавлять рост и развитие микроорганизмов.
Полная химическая структура установлена только для трети антибиотиков, а может быть получена химическим путем лишь половина из них. Синтез микроорганизмами антибиотиков - одна из форм проявления антагонизма, связан с определенным характером обмена веществ, возникшим и закрепленным в ходе эволюции. Воздействуя на постороннюю микробную клетку, антибиотик вызывает нарушения в её развитии.
В течение многих лет антибиотики используют как стимуляторы роста сельскохозяйственных животных и птицы, как средства борьбы с заболеваниями растений и посторонней микрофлорой в ряде бродильных производств, как консерванты пищевых продуктов. Механизм стимулирующего действия антибиотиков также не до конца выяснен. Предполагают, что стимулирующий эффект низких концентраций антибиотиков на организм животного связан с двумя факторами:
воздействие на микрофлору кишечника,
непосредственное влияние на организм животного.
В первом случае антибиотики снижают число вредных и увеличивают количество полезных для организма микроорганизмов. Во втором случае - снижают рН содержимого кишечника, уменьшают поверхностное натяжение клеток организма, что способствует ускорению их деления. Кроме того, антибиотики увеличивают количество ростовых гормонов, приспособляемость организма к неблагоприятным условиям и т.д. Кормовые антибиотики применяют в виде неочищенных препаратов, представляющих собой высушенную массу продуцента, содержащую помимо антибиотика аминокислоты, ферменты, витамина группы В и другие биологически активные вещества.
Все производимые кормовые антибиотики:
не используются в терапевтических целях и не вызывают перекрестной резистентности бактерий к антибиотикам, применяемым в медицине;
практически не всасываются в кровь из пищеварительного тракта;
не меняют своей структуры в организме;
не обладают антигенной природой, способствующей возникновению аллергии.
В настоящее время выпускаются несколько видов кормовых антибиотиков: препараты на основе хлортетрациклина (биовит, кормовой биомицин), бацитрацин, гризин, гигромицин Б и др. Из этих препаратов только бацитрацин представляет собой высушенную культуральную жидкость, полученную в результате глубинного выращивания Bacilus licheniformis. Остальные антибиотики являются продуктами жизнедеятельности разных видов Actinomyces.
Антибиотики используют и как средство борьбы с различными фитопатогенами. Воздействие антибиотика сводится к замедлению роста и гибели фитопатогенных микроорганизмов, содержащихся в семенах и вегетативных органах растений. К таким антибиотикам относятся фитобактериомицин, трихотецин, полимицин.
Применение антибиотиков в пищевой промышленности позволяет снизить длительность термообработки продуктов питания при их консервировании. Используемые антибиотики воздействуют на клостридиальные и термофильные бактерии, устойчивые к нагреванию. Наиболее эффективным признан низин, который практически не токсичен для человека и позволяет вдвое снизить время термообработки.
Микроорганизмы-фитостимуляторы
Задание «Разработать и внедрить высоко-эффективный полифункциональный препарат на основе генетически измененных ризосферных бактерий для защиты сельскохозяйственных культур от болезней различной этиологии и стимуляции их роста» (2007-2011 гг.)
Использование микроорганизмов в животноводстве
В настоящее время микроорганизмы в животноводстве используются в качестве биологических регуляторов метаболических процессов и как безопасная альтернатива низким дозам антибиотиков для предотвращения и лечения желудочно-кишечных расстройств у сельскохозяйственных животных (пробиотиков, эубиотиков, пребиотиков, симбиотиков, ферментных препаратов и витаминов).
Пробиотики – средства, восстанавливающие микробиоценозы. Согласно определению ВОЗ/FAO, пробиотики — это живые микроорганизмы, применённые в адекватных количествах, оказывающие оздоровительный эффект на организм человека (и животных).
Эубиотики – это бактерийные препараты, действующим началом которых являются живые лиофильно высушенные культуры микроорганизмов.
Пребиотики – это вещества, активизирующие рост нормальной кишечной микрофлоры.
Симбиотики представляют собой сочетание про- и пребиотиков, что потенциально улучшает выживаемость и приживаемость в кишечнике пробитиков.
Значение грибов в хозяйственной деятельности человека. Грибы и здоровье: подразделение грибных организмов с точки зрения медицинской микологии
Грибы имеют большое значение в природе и хозяйственной деятельности человека. Некоторые из них участвуют в круговороте веществ, разлагая растительные остатки и пополняя запас минеральных веществ в почве. Их ещё называют грибы-сапрофиты.
К сапрофитам можно отнести и дрожжи. Они развиваются на сахаристой среде и вызывают спиртовое брожение. Их широко используют в кулинарии, виноделии, для получения технического спирта. Пивные дрожжи продают в аптеках и используют для устранения гиповитаминоза, так как в их состав входят тиамин, рибофлавин, никотиновая кислота и другие витамины. Пищевые дрожжи на более чем на 50% состоят из белка, который по своему составу близок к белку мяса. В сельском хозяйстве интенсивно используются кормовые дрожжи.
Пеницилл с успехом применяется в пищевой промышленности. Его поселяют на некоторых сортах сыров с целью придания им специфического вкуса и аромата.
Как продукт питания, гриб используется уже многие тысячелетия. Всего насчитывается около 200 видов съедобных грибов. Шляпочные грибы содержат большое количество солей и витаминов. Белки содержатся в таких грибах в количестве около 30%. Организм человека усваивает только 2/3 от этого количества.
Существуют и грибы-паразиты, которые селятся на всех частях растений. Иногда поражаются культурные растения, что существенно снижает их урожайность. Например, на злаковых часто поселяются спорынья и головня. А вот картофель чаще всего страдает от фитофторы. Грибы могут поражать овощи и фрукты («мучнистая роса», «чёрная гниль»). Разрушают древесину домовой гриб, а также трутовик. Часто споры трутовика попадают в толщу дерева через повреждения. Гриб разрастается, делая древесину негодной, трухлявой.
Грибы и здоровье: медицинская микология
Грибы широко распространены в природе. Однако только небольшая часть их патогенна для животных или человека. Они обитают в почве, на растениях, паразитируют на животных и человеке. По характеру патогенности грибы подразделяются на монопатогенные, вызывающие заболевание только у животных (зоофильные) или у человека (антропофильные), и полипатогенные, способные поражать тех и других, т.е. зооантропофильные и антропозоофильные. Патогенные для человека грибы, поражающие кожу, называют еще дерматофитами, а заболевания - дерматомикозами.
Основную группу патогенных грибов составляют низшие растительные микроорганизмы, образующие ветвящиеся двухконтурные нити мицелия (длиной от 4 до 40-50 мкм и более, толщиной от 1 до 6 мм) и размножающиеся спорами. Они паразитируют в почве, на растениях, у животных и человека. К группе антропофильных грибов относят, например, фиолетовый и кратеровидный трихофитоны - возбудители поверхностной и хронической трихофитии, ржавый микроспорум, вызывающий «ржавую» микроспорию, ахорион Шенлейна - возбудитель фавуса, к группе зооантропофильных грибов - гипсовидный и фавиформный трихофитоны, вызывающие у телят, мышей и других животных трихофитию, а у человека ее инфильтративно-нагноительную форму, пушистый микроспорум, вызывающий заболевание как у собак и кошек, так и у человека.
Особую группу составляют дрожжеподобные патогенные грибы рода Кандида. Их основное отличие заключается в том, что они не образуют спор, размножаются почкованием, а образуемые ими нити не имеют настоящего ветвления, и поэтому их называют псевдомицелием.
Вирулентность грибов зависит от почвы, состояния растений, характера кожного покрова человека и животных. Инфицирование происходит двумя путями. Прямой путь передачи инфекции имеет место при контакте с почвой, растениями, больными животными или больным человеком, непрямой - при соприкосновении с различными вещами и предметами, бывшими в употреблении у больных, а также через предметы ухода за животными.
Грибковые инфекции являются важной проблемой клинической медицины. Из потенциальных "болезней будущего" микозы превратились в актуальные "болезни настоящего". Возбудители микозов многочисленны и вызываемые ими заболевания человека и животных весьма разнообразны. Количество видов микроскопических грибов (микромицетов) оценивается ~ 100000–200000 видов. Ежегодно описывается ~1500 новых видов. Реальное значение в клинической практике в настоящее время имеют около 100 видов.
Грибковые инфекции можно подразделить на контагиозные и оппортунистические. Контагиозные микозы часто эндемичны. Так, кокцидиоидоз характерен для стран с жарким сухим климатом (Центральная Америка, Мексика, Калифорния, Аргентина). Гистоплазмоз, вызываемый Histoplasma duboisii, эндемичен для западной и центральной Африки. Риноспоридиоз - для Индии и Цейлона. Североамериканский бластомикоз (болезнь Джилкрайста) и Южноамериканский бластомикоз (паракокцидиоидоз) названы по месту обнаружения. В России и странах СНГ эндемические зоны для этих заболеваний не обнаружены, хотя неоднократно отмечались случаи "завоза" их из-за рубежа.
Особое значение в последние десятилетия приобрели так называемые оппортунистические инфекции. Такие заболевания развиваются на фоне разнообразных иммуносупрессивных состояний. Загрязнение окружающей среды, повышение радиационного фона, использование в медицине иммуносупрессантов, цитостатиков, кортикостероидов, антибиотиков широкого спектра действия – вот далеко не полный перечень факторов, ослабляющих естественные защитные механизмы человека и способствующих возникновению оппортунистических инфекций, вызываемых условно-патогенными грибами.
В эпоху появления и широкого распространения ВИЧ-инфекции, вызывающей прогрессирующий иммунодефицит и создающего благоприятные условия для развития оппортунистических инфекций, отмечено резкое увеличение частоты микозов, некоторые из которых являются непосредственной причиной смерти больных.
По результатам комплексного обследования ВИЧ-инфицированных больных было установлено, что микозы составляют 70% в структуре клинических проявлений ВИЧ-инфекции наряду с другими инфекционными осложнениями. 60-90% больных ВИЧ-инфекцией страдают различными формами кандидоза, 5-20% больных погибает от криптококкового менингита, у 60% больных диагностируют пневмоцистную пневмонию, нередки случаи гистоплазмоза и аспергиллеза.
Второй глобальной проблемой является прогрессивное увеличение частоты грибковых инфекций, прежде всего – инвазивных микозов, у больных с различными новообразованиями. Основной причиной этого является широкое применение интенсивной цитостатической терапии, способствующей развитию нейтропении и дефектам клеточного иммунитета. Возможности полихимиотерапии опухолей и гемобластозов позволяют продлить жизнь больных, но в то же время у 50% этих больных обнаруживают микотические осложнения.
Очень высока летальность, которая составляет 25-55% при инвазивном кандидозе и 45-100% - при инвазивном аспергиллезе. Для некоторых категорий больных (реципиенты трансплантатов, острый лейкоз) инвазивные микозы являются основной причиной летального исхода.
Еще одна важная проблема связана с купированием внутрибольничных грибковых инфекций. В последние десятилетия отмечены вспышки внутрибольничных инфекций в самых совершенных, технически оснащенных больницах и клиниках. Особенно это касается отделений недоношенных детей, а также реанимационных отделений.
Интересы медицинской микологии связаны не только с заболеваниями, вызванными истинными грибами (эумицетами), но и с патологией, связанной с некоторыми водорослями класса Oomycetes. Так, водоросль Pythium insidiosum является возбудителем поражений подкожной клетчатки – фитиоза, а бесхлорофильный мутант зеленых водорослей Prototheca spp. Вызывает веррукозное поражение кожи и диссеминированные процессы с вовлечением лимфатических узлов и внутренних органов.
Медицинская микология традиционно занимается также патогенными актиномицетами, хотя эти микроорганизмы не относятся к грибам. Актиномицеты являются прокариотами (бактериями), содержат в стенках характерную мураминовую кислоту, у них отсутствуют оформленные ядра, они чувствительны к противобактериальным препаратам. Однако в культуре и тканях человека актиномицеты образуют тонкий ветвящийся мицелий, в связи с чем эти микроорганизмы длительное время относили к грибам. В настоящее время заболевания, вызываемые актиномицетами и нокардиями, называют псевдомикозами.
Билет № 6
Вирусы и бактерии как возбудители заболеваний. Наиболее известные вирусные и бактериальные болезни человека.
Патогенными называются бактерии, паразитирующие на других организмах. Бактерии вызывают большое количество заболеваний человека, таких как чума (Yersinia pestis), сибирская язва (Bacillus anthracis), лепра (проказа, возбудитель: Mycobacterium leprae), дифтерия (Corynebacterium diphtheriae), сифилис (Treponema pallidum), холера (Vibrio cholerae), туберкулёз (Mycobacterium tuberculosis), листериоз (Listeria monocytogenes) и др. Открытие патогенных свойств у бактерий продолжается: в 1976 г. обнаружена болезнь легионеров, вызываемая Legionella pneumophila, в 1980—1990 гг. было показано, что Helicobacter pylori вызывает язвенную болезнь и даже рак желудка, а также хронический гастрит.
Бактериальным инфекциям подвержены также растения и животные.
Многие бактерии, являющиеся в норме безопасными для человека или даже обычными обитателями его кожи или кишечника, в случае нарушения иммунитета или общего ослабления организма могут выступать в качестве патогенов.
Опасность бактериальных заболеваний была сильно снижена в конце XIX века с изобретением метода вакцинации, а в середине XX века с открытием антибиотиков.
Бактерии, вызывающие болезни человека, используются как биологическое оружие.
Различают две стратегии паразитизма. Первая стратегия создает преимущества паразиту в высокоплотных популяциях хозяев. Вторая, наоборот, оптимальна для паразитирования в редких, разбросанных на больших территориях, популяциях хозяев.
Современные представления о факторах патогенности болезнетворных бактерий
Инфекция – сложный биологический процесс, возникающий в результате проникновения патогенных микробов в организм и нарушения постоянства его внутренней среды. Возникновение инфекции зависит от нескольких факторов: степени патогенности (вирулентности) микроба, состояния макроорганизма и условий внешней среды.
Патогенность – это способность микроба определенного вида при соответствующих условиях вызывать характерное для него инфекционное заболевание. Следовательно, патогенность есть видовой признак.
Вирулентность – это степень патогенности определенного штамма микроба, т. е. индивидуальный признак. Например, бацилла сибирской язвы является патогенной, так как обладает свойством вызывает заболевание сибирской язвой. Но штамм одной культуры вызывает заболевание и смерть через 96 часов, а другой – через 6-7 дней. Следовательно, вирулентность первого штамма более высокая, чем второго.
Патогенность как особое качество болезнетворного вида микроба проявляется в агрессивных его свойствах и в токсическом действии на организм.
Агрессивность – это способность патогенного микроба жить, размножаться и распространяться в организме, противостоять неблагоприятным влияниям, оказываемым организмом. Некоторые патогенные микробы, размножаясь в организме или на питательной среде в пробирке, вырабатывают растворимые продукты, получившие название агрессины. Назначение агрессинов - подавлять действие фагоцитов. Сами агрессины безвредны для организма, но если их прибавить к несмертельной дозе культуры соответствующего микроба, они вызывают смертельно протекающую инфекцию.
Токсичность – способность патогенного микроба вырабатывать и выделять ядовитые вещества, вредно действующие на организм. Токсины бывают двух видов – экзотоксины и эндотоксины.
Экзотоксины – выделяются в окружающую среду при жизни микробов в организме или на искусственных питательных средах, а также в пищевых продуктах. Они очень ядовиты. Например, 0,005 мл жидкого столбнячного токсина или 0,0000001 мл ботулинического токсина убивает морскую свинку. Микробы, способные образовывать токсины, получили название токсигенных. Под влиянием нагревания и света экзотоксины легко разрушаются, а под действием некоторых химических веществ теряют токсичность.
Эндотоксины прочно связаны с телом микробной клетки и освобождаются только после ее гибели и разрушения. Они весьма устойчивы при действии высоких температур и не разрушаются даже после нескольких часов кипячения. Ядовитое действие многих бактерийных экзотоксинов связано с ферментами – лецитиназой (разрушает эритроциты), коллагеназой, гиалуронидазой (расщепляет гиалуроновую кислоту) и рядом других ферментов, которые производят в организме разрушение жизненно важных соединений. Установлено также, что некоторые патогенные бактерии (дифтерийные стафилококки и стрептококки) продуцируют фермент дезоксирибонуклеазу. В процессе жизнедеятельности патогенные микробы выделяют и другие вещества, обусловливающие их вирулентность.
Место проникновения патогенных микробов в организм называется входными воротами инфекции. В естественных условиях заражение происходит через пищеварительный тракт (алиментарный путь), когда в пищу или в воду попадают патогенные микроорганизмы. Болезнетворное начало может проникать через поврежденные, а при некоторых инфекционных болезнях (бруцеллез) и неповрежденные слизистые оболочки рта, носа, глаз, мочеполовых путей и кожу.
Судьба патогенных микробов, попавших в организм, может быть различной – в зависимости от состояния организма и вирулентности возбудителя.
Некоторые микробы, попав с током крови в определенные органы, оседают (задерживаются) в их тканях, размножаются в них, выделяют токсины и вызывают заболевание. Пример – возбудитель туберкулеза в легочной ткани.
Если патогенные микробы проникли в кровеносные сосуды и начинают размножаться в крови, то они очень быстро проникают во все внутренние органы и ткани. Такую форму инфекции называют септицемией. Она характеризуется быстротой и злокачественностью течения и нередко заканчивается смертельным исходом.
Когда микробы находятся в крови временно и не размножаются в ней, а посредством ее только переносятся в другие чувствительные ткани и органы, где затем уже размножаются, инфекцию принято называть бактериемией.
Иногда микробы, проникнув в организм, остаются только в поврежденной ткани и, размножаясь выделяют токсины. Последние, проникая в кровь, вызывают общее тяжелое отравление (столбняк, злокачественный отек). Такой процесс называется токсемией.
Пути выделения патогенных микробов из организма также различны: со слюной, мокротой, мочой, калом, молоком, выделениями из родовых путей.
Факторы патогенности отвечают за проявление конкретных свойств микроорганизма в инфекционном процессе. К ним относят:
факторы адгезии и колонизации — с их помощью бактерии распознают рецепторы на мембранах клеток, прикрепляются к ним и колонизируют клетки (различные поверхностные структуры клеточной стенки);
факторы инвазии — благодаря им бактерия проникает в клетку (белки наружной мембраны);
факторы, препятствующие фагоцитозу — либо маскируют бактерию от фагоцитоза (капсула), либо подавляют фагоцитоз (различные белки — белок А у стафилококков, белок М у стрептококков);
факторы, подавляющие фагоцитоз — вещества, подавляющие окислительный взрыв фагоцитов (например, V-W-антигены Y. pestis);
ферменты «защиты и агрессии» бактерий — способствуют распространению бактерий по тканям хозяина (гиалуронидаза, лецитиназа, протеазы и др.);
эндотоксины — представлены только у грамотрицательных микроорганизмов (липосахариды и связанные с ними белки клеточной стенки). Высвобождаются в среду организма после гибели клетки и обладают многообразным воспалительным и пирогенным действием неспецифического характера;
экзотоксины — токсические молекулы, активно секретируемые в окружающую среду с помощью специальных секретируемых систем.
Поскольку геномы бактерий небольшие (от 100 тыс до 4 млн пар нуклеотидов), многое удалось уже сделать в области функциональной геномики. И структурные, и функциональные исследования геномов патогенных бактерий показывают их высокую пластичность. Эти представления имеют непосредственное практическое значение:
во-первых, для разработки экспресс-методов типирования бактерий и оценки риска бактериальной контаминации;
во-вторых, для создания лекарств, нацеленных на специфические мишени, блокирующие работу генов патогенности;
в-третьих, для более целенаправленного создания вакцин.
Патогенные (болезнетворные) вирусы
Вирусы для человека и животных являются облигатными паразитами. Внешним проявлением вирулентности вируса являются разрушение инфицированных вирусом клеток и возникающие в результате разрушения тканей физиологические изменения в организме хозяина. Они используют обе стратегии паразитизма в самых разнообразных проявлениях. Мы ограничимся сравнением патогенных свойств двух, крайне опасных для людей вирусов — ВНО и ВИЧ. Первый из них (ВНО – вирус натуральной оспы) уже унес десятки миллионов жизней, второму, видимо, еще предстоит это сделать. Эти вирусы используют альтернативные стратегии паразитизма, у них совершенно несходные пути передачи, эпидемиология, патогенез и многое другое. Однако их сближает удивительная адаптированность к человеку, необычайно высокая способность распространяться в человеческих популяциях, продолжительность поддержания эпидемических очагов без постоянной циркуляции в природных резервуарах и, еще, видимо, древность, то есть способность появляться как бы ни откуда, собирать свою смертельную жатву, а затем исчезать на столь длительное время, что для потомков людей, переживших вызванные ими эпидемии они становятся возбудителями новых болезней.
Жизненный цикл ВИЧ. Связывание вириона с мембраной клетки (рецепция) возможно только при слиянии вирусного gp120 со специфическим рецептором на мембране клетки хозяина. После проникновения вириона, происходит его «раздевание», когда вследствие точечного протеолиза освобождается вирусная РНК. Центральным событием является синтез ДНК на матрице РНК. Уникальный фермент ретровирусов — обратная транскриптаза или ревертаза — разрушает нити РНК и синтезирует новую нуклеиновую кислоту, ДНК. Зрелые вирионы, образовавшиеся в клетке хозяина, отпочковываются, разрушая при этом клетку. Очень существенно, что репликация вируса с образованием множества вирионов, происходит только тогда, когда инфицированная клетка хозяина находится в активированном состоянии. В неактивированной клетке ВИЧ может находиться в латентном состоянии (в стадии провируса) неопределенно долго и с этим связана его кардинальная особенность его эпидемиологии как медленного вируса.
Происхождение вирусов
Вирусы являются фрагментами генома, способными к репликации только в пределах интактной живой клетки, следовательно, они не могут быть предшественниками клеток.
В пределах взятого вида, вирусы могут возникать как генетические фрагменты или редуцированные версии хромосом, плазмид или РНК:
1) хозяина или близкородственных видов;
2) отдаленных видов;
3) более крупных паразитов этого или другого хозяина;
4) дальнейшей эволюции или генетического обмена между существующими вирусами.
В прошлом они уже могли иметь жизненный цикл в геноме хозяина как интегрирующаяся эписома. В геноме они могут выполнять определенные генетические функции и, в принципе, способны вновь появляться в виде новых вирусов.
Существование таких циклов показано в мире бактериальных вирусов, однако мы не имеем четких данных, объясняющих происхождение вирусов животных и растений.
Общие принципы вирусного патогенеза. Для того, чтобы быть патогенными, вирусы, так же как и бактерии, должны:
инфицировать слизистые поверхности;
проникать в хозяина через эти поверхности; вступать в контакт с чувствительными тканями;
размножаться во внутренней среде хозяина;
противодействовать его защитным механизмам и причинять ему вред.
Необходимость в выполнении первых двух требований отпадает, если вирусы проникают в ткани через альвеолы (в составе мелкодисперсного аэрозоля), поврежденную переносчиком кожу или через слизистые.
Общие принципы вирусного патогенеза представлены в упрощенной схеме в следующей таблице. В правом столбце таблицы перечислены события, происходящие внутри клетки с момента начала ее инфицирования. Эти внутриклеточные стадии репликации вирусов, в конечном счете, определяют судьбу инфицированной клетки, т.е. ведут к ее гибели; клеточной трансформации, характеризующейся снятием ограничения роста; персистентной или латентной инфекции. В левом столбце приведены события на уровне организма или ткани, ведущие к инфицированию клеток и включающие иммунный ответ хозяина.
Хозяйские и клеточные стадии вирусного патогенеза
Организм |
Клетка |
Проникновение вируса в хозяина |
Адсорбция |
Первичная репликация |
Проникновение |
Распространение вируса внутри хозяина |
Раздевание |
Клеточный и тканевой тропизм и клеточные рецепторы |
Транскрипция |
Повреждение клеток |
Трансляция |
Имунный ответ и другие защитные факторы хозяина |
Сборка вируса |
Персистенция вируса, летентность и межденные вирусные инфекции |
Выход из клетки |
Вирусы, так же как и бактерии, должны быть способными преодолеть слизистые барьеры и проникнуть в те органы и ткани, где происходит их репликация. Прежде чем распространиться по организму, они могут реплицироваться в месте первичного проникновения и в ближайших регионарных лимфатических узлах. В отличие от бактерий, вирусы могут использовать нейрональный путь проникновения в поражаемые ими органы-мишени (вирус бешенства) и тем самым избежать противодействия иммунной системы. Однако чаще всего они, как и бактерии, распространяются с кровью и лимфой (вирусемия). С наибольшим постоянством вирусемия наблюдается при острых инфекциях, особенно тяжелых формах болезни. Однако циркуляция вирусов в крови установлена и при многих латентных, хронических инфекциях. Вирусы, свободно циркулирующие в плазме, представляют плазменный тип вирусемии (вирус Синдбис, ВЭЛ, японского энцефалита). Более распространенным является клеточный тип вирусемии, при котором вирусы циркулируют в тесной связи с клетками крови.
ВНО циркулируют, адсорбируясь на лимфоцитах и моноцитах крови, возбудители кори, гриппа, аденовирусы тесно связываются с полиморфно-ядерными лейкоцитами. Специфическое сродство вирусов к клеткам и тканям определяется присутствием на клеточной поверхности особых рецепторов. Отдельные; вирусы, после проникновения в клетку, могут индуцировать образование на ее поверхности дополнительных рецепторов, которыми «пользуются» другие вирусы. Метаболизм вирусов направлен на использование структур цитоплазматического или ядерного белкового скелета клеток. Вирусспецифический цитопатический эффект обусловлен не бессмысленным для вируса повреждением клетки, а специфической перестройкой элементов клеточного скелета, цель которой — создание условий для размножения вируса. Основным внешним проявление вирулентности вируса, являются разрушение инфицированных вирусом клеток в тканях-мишенях и возникающие в результате разрушения тканей физиологические изменения в организме хозяина.
У иммунокомпетентного хозяина большинство вирусных инфекций, возбудители которых используют первую стратегию паразитизма, самоограничиваются. Смертельные исходы в эпидемических очагах могут достигать очень высоких значений, однако в крупных популяциях людей они никогда не достигают 100%. При недостаточности иммунной системы хозяина, эти же вирусы могут вызывать персистентные или латентные инфекции.
Персистирующая инфекция. Это бессимптомная персистенция вируса, сопровождающаяся его выделением в окружающую среду. В механизме развития вирусной персистирующей инфекции значительная роль отводится особому состоянию возбудителя — дефектным интерферирующим частицам вируса или его мутантам, а также блокированию апоптоза клеток хозяина. Персистирующий инфекционный процесс лежит в основе латентной инфекции.
Латентная инфекция. Это бессимптомная персистенция вируса, не сопровождающаяся его выделением в окружающую среду. При латентной инфекции вирус не удается обнаружить с помощью диагностических приемов в связи с тем, что он находится в дефектной форме или интегрирован в геном клетки. Этот процесс плохо изучен. Значительно большая ясность имеется в вопросе реактивации вируса. Считается, что для этого необходимо действие одного или нескольких активирующих факторов (травма, стресс, иммуносупрессия, суперинфицирование и др.). В результате латентная инфекция переходит в манифестную форму — развиваются симптомы, свойственные острой инфекции. Примерами латентных инфекций служат герпесвирусные, цитомегаловирусные инфекции, сывороточные гепатиты и ряд других. Способность вызывать персистирующую или латентную инфекцию не является общим свойством для всех вирусов. Например, ВНО и отдельные возбудители геморрагических лихорадок либо вызывают смерть инфицированного, либо разрешаются с развитием стерильного иммунитета. Сколь бы многочисленной не осталась пережившая эпидемию популяция, они в ней не сохраняются, а для их поддержания в природе используются другие механизмы.
Что касается геномики вирусов, то для большинства патогенных для человека вирусов (возбудителей вирусных гепатитов, ВИЧ-инфекции и СПИДа, герпесвирусных инфекций, натуральной оспы, гриппа и др.) уже известна первичная нуклеотидная последовательность полноразмерного генома (структурная геномика). Более того, накоплено много данных по функциональной геномике (роль отдельных фрагментов в формировании вторичной структуры генома, в образовании белков вирионов, в репликации и сборке вирионов). Именно геномные исследования вирусов позволили объяснить их высокую пластичность (способность к рекомбинации, наличие гипервариабельных областей). Многие вирусы формируют длительную персистентную инфекцию, в результате которой происходит селекция новых вариантов вируса с изменённой первичной последовательностью, а следовательно, с изменёнными патогенными и антигенными свойствами.
Несмотря на интенсивные поиски участков в геномах вирусов (сайтов), ответственных за патогенные свойства вирусов, они до сих пор не обнаружены, т.е. функциональная геномика вирусов ещё не достигла такого уровня, как структурная. Результаты исследований позволяют с большой вероятностью думать о том, что патогенные свойства вирусов являются полифункциональным признаком, детерминируемым многими сайтами генома.
Практическое приложение сведений о нуклеотидной последовательности геномов многих патогенных вирусов уже широко реализуется. Генно-инженерным путём создаются непатогенные фрагменты геномов вирусов в составе плазмидных векторов. Такие векторы с вирусом способны к экспрессии в высоких концентрациях белков вирусов, которые необходимы для приготовления диагностических и вакцинных препаратов. Развивается технология получения ДНК-вакцин против СПИДа, гепатита С и других вирусных инфекций. Создана эффективная рекомбинантная вакцина против гепатита В.
Природные ресурсы: их структура, рациональное использование и охрана.
ПРИРОДНЫЕ РЕСУРСЫ (natural resources), естественные ресурсы — часть всей совокупности природных условий и важнейших компонентов природной среды, которые используются либо могут быть использованы для удовлетворения разнообразных потребностей общества и общественного производства. Условно подразделяются на неисчерпаемые, почерпаемые возобновимые и исчерпаемые невозобновимые, на реальные и потенциальные, на заменимые и незаменимые. По отношению к тем или иным компонентам природы различают группы и виды природных ресурсов: биологические, экологические, геологические, климатические, водные, земельные, минеральные, генетические, растительные, лесные, ресурсы животного мира и т. д. По характеру использования в производственной и непроизводственной сферах природные ресурсы классифицируют на промышленные, сельскохозяйственные, минерально-сырьевые, топливно-энергетические, продовольственные, оздоровительные, ландшафтно-курортные, рекреационные и др.
Все природные ресурсы по исчерпаемости делятся на две группы: исчерпаемые и неисчерпаемые.
I. Исчерпаемые ресурсы. Они образуются в земной коре или ландшафтной сфере, но объемы и скорости их формирования измеряются по геологической шкале времени. В то же время потребности в таких ресурсах со стороны производства или для организации благоприятных условий обитания человеческого общества значительно превышают объемы и скорости естественного восполнения. В результате неизбежно наступает истощение запасов природного ресурса. В группу исчерпаемых включены ресурсы с неодинаковыми скоростями и объемами формирования. Это позволяет провести их дополнительную дифференциацию.
На основе интенсивности и скорости естественного образования исчерпаемые ресурсы делят на подгруппы:
1. Невозобновляемые, к которым относят:
а) все виды минеральных ресурсов или полезные ископаемые. Они постоянно образуются в недрах земной коры в результате непрерывно протекающего процесса рудообразования, но масштабы их накопления столь незначительны, а скорости образования измеряются многими десятками и сотнями миллионов лет (например, возраст каменных углей насчитывает более 350 млн. лет), что практически их учитывать в хозяйственных расчетах нельзя. Освоение минерального сырья происходит по исторической шкале времени и характеризуется всевозрастающими объемами изъятия. В этой связи все минеральные ресурсы рассматриваются в качестве не только исчерпаемых, но и невозобновляемых.
б) земельные ресурсы в их естественном природном виде - это материальный базис, на котором происходит жизнедеятельность человеческого общества. Морфологическое устройство поверхности (т. е. рельеф) существенно влияет на хозяйственную деятельность, на возможность освоения территории. Однажды нарушенные земли (например, карьерами) при крупном промышленном или гражданском строительстве в своем естественном виде уже не восстанавливаются.
2. Возобновляемые ресурсы, к которым принадлежат:
а) ресурсы растительного и
б) животного мира.
И те и другие восстанавливаются довольно быстро, и объемы естественного возобновления хорошо и точно рассчитываются. Поэтому при организации хозяйственного использования накопленных запасов древесины в лесах, травостоя на лугах или пастбищах, промысла диких животных в пределах, не превышающих ежегодное возобновление, можно полностью избежать истощения ресурсов.
3. Относительно (не полностью) возобновляемые. Некоторые ресурсы хотя и восстанавливаются в исторические отрезки времени, но возобновляемые объемы их значительно меньше объемов хозяйственного потребления. Именно поэтому такие виды ресурсов оказываются весьма уязвимыми и требуют особенно тщательного контроля со стороны человека. К относительно возобновляемым ресурсам относятся и очень дефицитные природные богатства:
а) продуктивные пахотно-пригодные почвы;
б) леса с древостоями спелого возраста;
в) водные ресурсы в региональном аспекте.
Продуктивных пахотно-пригодных почв сравнительно немного (по разным оценкам их площадь не превышает 1,5-2,5 млрд. га). Наиболее продуктивные почвы, относящиеся к первому классу плодородия, занимают, по оценкам ФАО, всего 400 млн. га. Продуктивные почвы образуются крайне медленно – на формирование 1 мм слоя, например, черноземных почв требуется более 100 лет.
II. Неисчерпаемые ресурсы. Среди тел и явлений природы ресурсного значения имеются и такие, которые практически неисчерпаемы, К ним относятся климатические и водные ресурсы.
А) Климатические ресурсы. Наиболее жесткие требования к климату предъявляют сельское хозяйство, рекреационное и лесное хозяйство, промышленное и гражданское строительство и др. Обычно под климатическими ресурсами понимают запасы тепла и влаги, которыми располагает конкретная местность или регион. Общие запасы тепла, поступающие за год на 1 м.кв. поверхности планеты, равны 3.16 х 10 Дж (радиационный бюджет в среднем для планеты). Территориально и по сезонам года тепло распределяется неравномерно, хотя в среднем для Земли температура воздуха равна примерно + 15°С. Суша в целом неплохо обеспечена и атмосферной влагой: на ее поверхность ежегодно выпадает в среднем около 119 тыс. куб. км осадков. Но распределяются они еще более неравномерно, чем тепло, и в пространственном и во временном отношениях На суше известны районы, получающие ежегодно более 12000 мм осадков, к обширные местности, где за год выпадает менее 50-100 мм. В среднем многолетнем выражении и запасы тепла и объемы выпадающей атмосферной влаги довольно постоянны, хотя от года к году могут наблюдаться существенные колебания в обеспечении территории теплом и влагой. Так как эти ресурсы формируются в определенных звеньях теплового и водного круговоротов, постоянно действующих над планетой в целом и над ее отдельными регионами, запасы тепла и влаги могут рассматриваться как неиссякаемые в определенных количественных пределах, точно установленных для каждого района.
Б) Водные ресурсы. Земля обладает колоссальным объемом воды - около 1,5 млрд. куб. км. Однако 98% этого объема составляют соленые воды Мирового океана, и только 28 млн. куб. км - пресные воды. Поскольку уже известны технологии опреснения соленых морских вод, воды Мирового океана и соленых озер можно рассматривать как потенциальные водные ресурсы, использование которых в будущем вполне возможно. Ежегодно возобновляемые запасы пресных вод не столь велики: по разным оценкам они колеблются от 41 до 45 тыс. куб. км (ресурсы полного речного стока). Мировое хозяйство расходует для своих нужд около 4-4,5 тыс. куб. км, что равно примерно 10% общего водозапаса, и, следовательно, при условии соблюдения принципов рационального водопользования эти ресурсы можно рассматривать как неисчерпаемые. Однако при нарушении этих принципов ситуация может резко обостриться, и даже в планетарном масштабе может ощущаться дефицит чистых пресных вод. А пока природная среда ежегодно "дарит" человечеству в 10 раз больше воды, чем ему нужно для удовлетворения самых разнообразных потребностей.
Ресурсы - все извлекаемое нами из природной среды для удовлетворения своих потребностей и желаний (чистый воздух, питьевая вода, нефть, железная руда, древесина, культурные растения). Большая часть ресурсов требует для своего употребления определенных условий, определенного развития технологии (например, нефть, никелевые руды, бокситы). Значит, все, что угодно может стать ресурсом при соответствующих технологии и потребностях.
Типы ресурсов:
исчерпаемые (возобновимые и невозобновимые);
неисчерпаемые (солнечная энергия, энергия ветра, приливов, геотермальная энергия и т.д. Невозобновимые ресурсы существуют в ограниченных количествах (запасах) в различных частях земной коры. Примерами являются медь, алюминий, уголь и нефть. Невозобновимые ресурсы считаются экономически истощенными, когда выработаны 80% их оцененных запасов. По достижении этого предела разведка, добыча и переработка остающихся запасов обходится дороже розничной цены. Некоторые виды невозобновимых ресурсов могут использоваться повторно или циклически, увеличивая, таким образом, потенциал использования ресурсов алюминия, меди, железа или, например, стекла.
Потенциально возобновимые - ресурсы, запасы которых хотя и могут быть истощены или загрязнены в результате слишком быстрого потребления, однако в нормальных условиях восстановятся в результате естественных процессов (деревья, пресные воды рек и озер, почвы, дикие животные).
Утилизация или рециркуляция - переработка ресурсов в новую продукцию (алюминиевые банки).
Повторное использование - использование ресурса в той же форме (стеклянная бутылка).
Уровень устойчивого потребления - наиболее высокая скорость, при которой возобновимые ресурсы могут быть использованы без снижения возможности их возобновления. При превышении уровня устойчивого потребления происходит деградация ресурса.
Охрана ресурсов заключается в использовании, управлении и защите ресурсов, с тем чтобы ими могли пользоваться в устойчивом режиме как настоящее, так и будущее поколения.
Абсолютная недостаточность ресурсов возникает, когда нет достаточных или доступных запасов ресурсов для удовлетворения имеющихся или будущих потребностей.
Относительная недостаточность ресурсов возникает тогда, когда ресурсов еще хватает для удовлетворения потребностей, но их распределение не сбалансировано.
Под термином «БИОЛОГИЧЕСКИЕ РЕСУРСЫ» обычно подразумевается совокупность организмов, которые могут быть использованы человеком прямо или косвенно для потребления (лесные, растительные, рыбные, охотничьи и др.).
Биоресурсы – исчерпаемый возобновимый тип природных ресурсов. При оптимальном природопользовании в течение длительного времени численность и биомасса организмов, эксплуатируемых человеком, не снижаются и не изменяется структура их сообществ.
Биоресурсы суши
Биоресурсы растительного происхождения
Сосудистые растения
Мохообразные
Водоросли
Лишайники
Грибы
Биоресурсы животного происхождения
Позвоночные животные
Млекопитающие
Птицы
Рептилии
Амфибии
Рыбы
Круглоротые
Беспозвоночные животные
Водные биоресурсы
Вне этой классификации находятся:
Видовое и ценотическое разнообразие растительного мира
Лекарственно-техническое и пищевое дикорастущее сырье
Охотничьи ресурсы
Виды, охраняемые в искусственных условиях
Вопрос объективной оценки ресурсной значимости видов флоры достаточно сложен и не вполне разработан в методическом отношении. Во-первых, это связано с отсутствием официальной классификации хозяйственно полезных растений и четко определенных ресурсных групп. Во-вторых, каждый вид растений обладает потенциалом хозяйственного использования, имеет то или иное экологическое или информационное значение. В-третьих, многие виды растений используются в различных отраслях народного хозяйства в различном качестве, т. е. виды в большинстве своем полифункциональны в практическом отношении и могут входить в состав разных ресурсных групп и подгрупп (пищевых, лекарственных и прочих). По принципу возможного практического использования растений в народном хозяйстве и в быту выделены 10 ресурсных групп. Некоторые из них имеют подгруппы, в которых учтены особенности использования хозяйственно полезных видов (по целевому назначению и воздействию, по используемым частям, веществам и т. п.).
Экологически грамотная эксплуатация и охрана растительных ресурсов
Охрана растительного мира в Беларуси: 39 видов растений, произрастающих в Беларуси, имеют статус охраняемых в Европе. Эти виды включены в Бернскую конвенцию (19 видов), в Приложении IIb к Директиве Европейского союза по местообитаниям (19), в Приложение IVb к Директиве Европейского Союза по местообитаниям (22), в европейский список охраняемых макролишайников (2 вида) и в Красную книгу мохообразных Европы (4 вида). Среди них – 25 видов сосудистых растений, 6 – мохообразных, 2 вида лишайников и 6 видов грибов. В нашей стране отмечено 33 типа местообитаний, подлежащих охране в Европе согласно приложению к директиве по местообитаниям и 26 – согласно приложению к Бернской конвенции.
В соответствии с Программой «Зеленые легкие Европы», которая разработана министрами окружающей среды Беларуси, Латвии, Литвы, Польши, Российской Федерации, Украины и Эстонии и принята в феврале 1993 г., вся территория Беларуси включена в зону, которая представляет собой особую экологическую ценность для Восточной и Центральной Европы. В пределах зоны «Зеленых легких Европы» выделено 21 крупное природное территориальное образование, включающее наиболее ценные в природном отношении местности. 10 из них полностью или частично расположены на территории Беларуси.
Браславские озера (300 тыс. га); Белорусские озера, включая озеро Нарочь и Березинский биосферный заповедник (1730 тыс. га); Налибокская пуща (280 тыс. га); Бобруйско-Борисовский район (1260 тыс. га); Восточное Полесье; Беловежская пуща (960 тыс. га, в том числе в Беларуси 480 тыс. га); Литовские озера ; Витебские и Латгальские озера; Долина Западной Двины (2325 тыс. га, в том числе в Беларуси 255 тыс. га); Долина Буга.
Что касается заповедных и охраняемых территорий, то в них необходимо в будущем, используя опыт изучения и сохранения биоразнообразия в ГНП «Беловежская пуща» и Березинского биосферного заповедника провести полную инвентаризацию биоразнообразия различных групп растений и животных. Для этого необходимо по специально разработанным проектам для каждого из имеющихся в стране заповедников и заказников сконцентрировать на ключевых вопросах изучения биоразнообразия в следующих целевых междисциплинарных направлениях:
Инвентаризация и классификация биоразнообразия - оценка современного состояния биоразнообразия для проведения инвентаризации различных отдельных видов и их таксономических групп; анализ и синтез полученной информации будет использован для подготовки прогнозов дальнейших исследований и подготовки узкоспециализированных специалистов-систематиков.
Функционирование экосистем и поддержание биоразнообразия - получение ответов о стабильности экосистем и их способности к восстановлению в зависимости от разнообразия видов, о роли глобальных изменений климата, землепользования и техногенеза на функционирование экосистем, о роли биоразнообразия (от генетического и видового до ландшафтного уровней) в экосистемных процессах (изменение продуктивности, обеспечение декомпозиции и круговорота питательных веществ и т.д.).
Вопросы эволюции, происхождения и сохранения биоразнообразия - изучение реакции видов и различных хозяйственных групп растений на изменение среды их обитания с учетом различных научных подходов, включая генетические, физиологические и молекулярно-биохимические свойства видов, межвидовые отношения и популяционные уровни в процессе эволюции; выяснение физиологических, генетических процессов, обеспечивающих механизмы формо- и видообразования и видовой деградации, ведущие соответственно к увеличению и утрате регионального континентального и мирового биотического богатства.
Мониторинг биоразнообразия - предполагает разработку эффективных и стандартных методов наблюдения за состоянием биоразнообразия (видов, популяций, экосистем) с целью выработки критериев для подготовки и принятия обоснованных решений на любом уровне управления, направленных на смягчения этих изменений.
Охрана, восстановление и устойчивое (рациональное) использование биоразнообразия - изучение факторов (природных и антропогенных) влияющих на биоразнообразие, его динамику и рациональное использование, разработка системы охраны биоразнообразия, разработка стратегий по восстановлению нарушенных экосистем, генетическим изменениям популяций в процессе их восстановления.
Билет № 7
Протеомика и современные проблемы белковой инженерии.
Протеомика — наука, основным предметом изучения которой являются белки и их взаимодействия в живых организмах, в том числе — в человеческом. Учёные, работающие в области протеомики, исследуют «производство» белков, их декомпозицию и замену белков внутри тела. Они также изучают как белки модифицируются после их синтеза в организме. Традиционно изучение белков являлось одним из разделов биохимии, но после определения структуры всей геномной ДНК человека и ряда других организмов, у исследователей белков появились новые методы, с которыми и связывают появление нового термина протеомика (от протеин и геномика). В частности, появились исчерпывающие базы данных о структуре всех белков человека, а также их протеолитических фрагментов, полученных в стандартных условиях. Это позволяет идентифицировать белки по молекулярной массе их протеолитических фрагментов, полученных в тех же условиях
Принципы и методы анализа протеома.
Методы протеомных исследований
Исследование состава белков:
- 2Д-электрофорез;
- высокоэффективная жидкостная хроматография (ВЭЖХ);
- масс-спектрометрия (MALDI, ESI).
Исследование структуры белков:
2Д-электрофорез
Протеомный анализ начинают с получения образцов биологического материала и экстракции из них белков. Затем полученную смесь белков с целью их первичной идентификации разделяют методом двумерного гель-электрофореза (2Д-электрофорез). Этот метод обеспечивает разделение белков по двум физико-химическим свойствам – электрическому заряду и массе. При осуществлении метода 2Д-электрофореза белки вначале разделяют в одном направлении – по заряду, а затем во втором направлении – по массе. В результате получают 2Д-электрофореграмму.
2D-электрофорез изоэлектрическое фокусирование
2 D-электрофорез
нативный электрофорез (native PAGE)
денатурирующий электрофорез (SDS-PAGE)
D-электрофорез
нативный электрофорез (native PAGE)
денатурирующий электрофорез (SDS-PAGE)
2D-электрофорез(SDS-PAGE)
Действуют детергентом SDS. Это разделяет комплексы и разворачивает белки. К ним прикрепляются SDS-молекулы, заряженные отрицательно. Количество прикрепленных пропорционально размеру, т.е. грубо говоря пропорционально массе. В горизонтальном направлении направлении прилагается поле, которое их разделяет пропорционально e/m, но это отношение всюду одинаково.В вертикальном направлении их смещение обратно-пропорционально масс из-за трения. Т.е. PAGE действует, как молекулярное сито.
Масс-спектрометрия
Значительно более детальная, чем с помощью 2Д-электрофореза, идентификация белков может быть достигнута методами масс-спектрометрии. В процессе осуществлении масс-спектрометрического анализа исследуемые белки фрагментируют до пептидов и подвергают ионизации.
Существует две основные модификаций метода ионизации пептидов: матриксная лазерно-десорбционная ионизация – MALDI (Matrics-Assisted Laser Desorbtion Ionisation) и электроспрейная ионизация – ESI (Electrospray Ionisation).
В модификации MALDI (Matrix-assisted laser desorption/ionization) анализируемый препарат смешивают с матрицей – донором протонов и формируют таким образом мишень действия лазерного излучения. Под действие ультрафиолетового лазерного импульса наносекундной длительности происходит испарение материала мишени и ионизация анализируемых пептидов.
С помощью времяпролётного (TOF – Time-Of-Flight) масс-анализатора ионизированные пептиды разделяются по параметру отношения массы к заряду (m/z). Разделение происходит вследствие того, что в TOF-анализаторе разогнанные электрическим полем ионизированные пептиды движутся в зависимости от массы с разными скоростями и, соответственно, в разное время достигают детектора, расположенного в конце бесполевого пространства. Измерив время пролета ионизированных пептидов, можно определить их массу.
ESI (electrospray ionization)
В модификации ESI электроспрейная ионизация достигается путем распыления с помощью высокого напряжения (до 20 кВ) растворов анализируемых пептидов, подающихся по тонкому капилляру. Ионизированные пептиды разделяются затем в ESI масс-спектрометре. По сравнению с MALDI, ESI имеет преимущество в относительной простоте и возможности сочетания с техникой разделения белков методами жидкостной хроматографии. Результаты масс-спектрометрических измерений представляются в виде спектра.
Структурная и функциональная протеомика.
Протеомика: Структурная, функциональная, регуляторная протеомика.
Структурная протеомика:
Цель структурной протеомики: установление структуры всех белков живого организма.
Задачи структурной протеомики:
- выделение и очистка белков протеома;
- идентификация белков протеома;
- определение первичной, вторичной, третичной структуры белков;
Функциональная протеомика:
Цель функциональной протеомики: определение функциональных свойств протеома.
Задачи функциональной протеомики:
- установление функций каждого из белков протеома;
- предсказание функциональной роли отдельных белков путем сопоставления их качественного и количественного состава в клетке на разных стадиях и в разных состояниях ее развития;
- анализ межбелковых взаимодействий;
- выяснение взаимосвязи между структурой и функцией белков;
- изучение механизмов функционирования белков.
Регуляторная протеомика:
Целью регуляторной протеомики является выяснение закономерностей регуляции активности протеома
Задачи регуляторной протеомики:
- расшифровка механизмов регуляции активности протеома;
- установление механизмов регуляции функций индивидуальных белков;
- выяснение механизмов регуляции межбелковых взаимодействий.
Прикладные аспекты протеомики.
Исследования в области протеомики в настоящее время имеют выраженную медицинскую направленность. Одной из важнейших задач протеомики является выявление специфичных для каждого конкретного заболевания изменений в протеоме. Получение таких научных результатов облегчит диагностику патологии, позволит выявлять заболевания на ранних, предшествующих клиническим проявлениям стадиях, даст возможность выбрать эффективную стратегию лечения патологии. С помощью протеомики можно установить диспропорцию белков в патологически измененных тканях. Например, протеомный сравнительный анализ позволит выяснить, какие именно отклонения в белковом составе клеток являются характерными для конкретной злокачественной опухоли. А на основе этих данных можно получить антитела, способные эффективно подавлять рост раковых клеток.
Протеомика позволит выявлять целевые протеины (мишени) и тем самым способствовать созданию новых высокоэффективных медикаментозных и диагностических средств нового поколения. Обладая знаниями в области протеомики, можно будет быстро создавать препараты нового поколения с точной, адресной доставкой. Ожидается, что созданные методами протеомики препараты будут лишены побочного действия.
Ещё один медицинский аспект протеомики касается различной восприимчивости людей к лекарственным препаратам. Выяснение специфики изменений протеома у больных будет способствовать решению и этой проблемы. Пока не ясно, сбудутся ли все надежды и ожидания, связанные с протеомикой, но уже сейчас протеомика объявлена одной из важнейших отраслей науки XXI века.
Вид как основной объект при изучении биоразнообразия. Концепции вида. Аллопатрическое и симпатрическое видообразование.
Существуют две основные концепции вида: типологическая и биологическая.
Первая основана целиком на морфологическом критерии: вид обладает вполне определенным типом или планом строения и все, что не укладывается в рамки этого типа, не есть данный вид. В основу разграничения видов кладутся морфологические отличия. Типологическая концепция с успехом применяется в минералогии, но совершенно неприемлема для биологических объектов, поскольку она рассматривает живые организмы как совокупность неодушевленных предметов.
Основа биологической концепции вида - критерий скрещиваемости. Вид - это группа особей, репродуктивно изолированная от других таких групп. Биологическая концепция вида представлена, в свою очередь, двумя концепциями: концепцией одномерного и многомерного видов.
Концепция одномерного вида рассматривает взаимоотношение двух популяций, сосуществующих друг с другом, не смешивающихся в одном и том же месте и в одно и то же время, то есть симпатрические и синхронные популяции. Симпатрические популяции имеют резкие разрывы, отражающие обособленность видов. Таким образом, если в типологической концепции вид устанавливается на основании субъективно выбранных различий, то в концепции одномерного вида используются морфологические отличия, отражающие репродуктивную изоляцию данного вида от других.
В концепции многомерного вида (или концепции скрещивающихся популяций) вид рассматривается как группа популяций, фактически или потенциально скрещивающихся друг с другом. Данная концепция рассматривает пространственно разобщенные, или аллопатрические, популяции. Разобщенные в пространстве и времени популяции могут значительно отличаться друг от друга, хотя они и не изолированы репродуктивно. В данном случае мы имеем дело с видом, который представлен многими различными аллопатрическими популяциями.
Концепция многомерного вида наиболее полно отражает положение вещей в природе, однако в практической деятельности этой концепцией пользоваться крайне трудно. Все-таки вид Линнея и Джона Рея (одномерный вид) до сих пор занимает господствующее положение в зоологической систематике, во всяком случае в систематике насекомых.
Вид – это единственная таксономическая категория, которой можно дать относительно точное определение. Вот некоторые из определений вида, базирующиеся на приведенных выше концепциях:
Вид – это группа особей, обладающих единственным в своём роде набором морфологических (структурных) и функциональных признаков, т.е. внешним видом, особенностями расположения органов и их работы и т.п.
Вид – это группа особей, способных, скрещиваясь между собой, давать плодовитое потомство.
Вид – это группа особей, сходных по генотипу (количеству, размеру и форме хромосом).
Вид – это группа особей, занимающих одну и ту же экологическую нишу.
Для установления видовой принадлежности недостаточно использовать только один критерий; как правило, только совокупность критериев правильно характеризует вид. Вид составляют одна или несколько популяций. Популяция – это группа свободно скрещивающихся особей, длительное время существующих на территории, обособленной относительно других групп особей того же вида. Именно популяция является основной единицей эволюции. Как правило, вид распространён на достаточно большой территории; условия внешней среды могут настолько сильно различаться в разных местообитаниях, что это приводит к появлению рас и подвидов с резко выраженными отличиями (например, цвет кожи у людей разных рас). В других случаях фенотип по какому-либо признаку может изменяться достаточно плавно; в этом случае говорят о существовании клин. У одного вида может быть несколько клин (каждая – по своему признаку), различающихся по направлению.
Как правило, вид распространён на достаточно большой территории; условия внешней среды могут настолько сильно различаться в разных местообитаниях, что это приводит к появлению рас и подвидов с резко выраженными отличиями (например, цвет кожи у людей разных рас). В других случаях фенотип по какому-либо признаку может изменяться достаточно плавно; в этом случае говорят о существовании клин. У одного вида может быть несколько клин (каждая – по своему признаку), различающихся по направлению.
Наличие нескольких генетически обособленных популяций может привести к видообразованию – возникновению нового вида. Если этот вид образуется из одного вида, существовавшего ранее, то говорят о внутривидовом видообразовании. Непременным его условием является прекращение обмена генами между популяциями. Внутривидовое видообразование может протекать в условиях пространственной разобщён-ности популяций (аллопатрическое видообразование) и в том случае, когда популяции занимают одну и ту же географическую область (симпатри-ческое видообразование).
П ри
аллотропическом
видообразовании
популяции должны долгое время находиться
на географически разобщённых территориях.
Ослабление конкурентного отбора,
адаптация к новым условиям и случайный
дрейф генов приводят к повышению
внутривидовой изменчивости; в результате
в генотипе популяции изменяются частоты
аллелей и генотипов. Если популяции
через какое-то время снова встречаются,
то они либо продолжают скрещиваться,
либо скрещивание между ними становится
невозможным; во втором случае образуются
новые виды.
ри
аллотропическом
видообразовании
популяции должны долгое время находиться
на географически разобщённых территориях.
Ослабление конкурентного отбора,
адаптация к новым условиям и случайный
дрейф генов приводят к повышению
внутривидовой изменчивости; в результате
в генотипе популяции изменяются частоты
аллелей и генотипов. Если популяции
через какое-то время снова встречаются,
то они либо продолжают скрещиваться,
либо скрещивание между ними становится
невозможным; во втором случае образуются
новые виды.
Симпатрическое видообразование происходит в тех популяциях, которые обитают на одной территории. Как правило, это связано с появлением в популяции нескольких групп, особи которых не могут скрещиваться между собой.
Факторы, мешающие скрещиванию особей из различных популяций, – изолирующие механизмы – могут быть следующими:
экологическая изоляция – особи различных групп предпочитают различные условия местообитания;
сезонная изоляция – оплодотворение у особей разных групп происходит в различное время;
поведенческая изоляция – особи разных групп имеют различные ритуалы ухаживания;
механическая изоляция – репродуктивные органы особей разных групп делают невозможной перекрёстное оплодотворение;
нежизнеспособность гибридов – гибриды погибают в раннем возрасте;
стерильность гибридов – гибриды первого поколения не способны производить функциональные гаметы (в частности, из-за наличия в диплоидной клетке нечётного числа хромосом);
неполноценность гибридов – гибриды первого поколения способны производить потомство, но следующее поколение либо неполноценно? либо стерильно.
Особый случай симпатрического видообразования наблюдается в случае кольцевых клин, если популяции последовательно друг за другом заселяют какую-либо географическую область, замыкая кольцо. Краевые популяции могут при этом оказаться генетически настолько различными, что будут неспособны образовывать плодовитое потомство.
Межвидовая гибридизация возникает, когда между собой скрещиваются особи разных видов. Плодовитое потомство, как правило, появляется при наличии хромосомных мутаций, в результате которых образуется аллополиплоид.
Образование новых видов и подвидов называется микроэволюцией в отличие от макроэволюции, приводящей к появлению или вымиранию целых таксономических групп организмов.
Билет № 8
Типы антропогенных экосистем и основные их характеристики.
Человек и экосистемы.
За последние 50 лет условия существования человека заметно ухудшились. И эта тенденция лишь нарастает. Чтобы доказать экологическую пропасть, в которую движется человечество, грядущую экологическую катастрофу, сегодня уже не нужно быть специалистом-экологом. Достаточно посмотреть и оценить, как и с какой скоростью изменились за последние сто или пятьдесят лет отдельные природные ресурсы или отдельные территории на планете Земля. Сколько было рыбы в реках и морях, ягод и грибов в лесах, цветов и бабочек на лугах, лягушек и птиц в болотах, зайцев и других пушных зверей и т.д. сто, пятьдесят, двадцать лет назад? Всё меньше, меньше, меньше… Подобная картина характерна для большинства групп животных, растений и отдельных неживых природных ресурсов. Красная книга исчезающих и становящихся редкими видов постоянно пополняется новыми жертвами деятельности человека "разумного"...
А качество и чистота воздуха, воды и почвы сто, пятьдесят лет назад и сегодня! Ведь там, где живет человек, сегодня кругом бытовые отходы, не разлагаемый в природе пластик, опасные химические выбросы, автомобильные выхлопные газы и прочие загрязнения. Леса вокруг городов, заваленные мусором, висящие над городами смоги, коптящие в небо трубы электростанций, фабрик и заводов, загрязненные или отравленные стоками реки, озера и моря, перенасыщенная удобрениями и ядохимикатами почва и грунтовые воды… А каких-то сто лет назад многие территории были почти девственны с точки зрения сохранности дикой природы и отсутствия там человека.
Широкомасштабная мелиорация и осушение, вырубка лесов, сельскохозяйственное освоение земель, опустынивание, строительство и урбанизация - территорий интенсивного хозяйственного использования становится всё больше, территорий дикой природы – всё меньше. Нарушается баланс, равновесие между дикой природой и человеком. Естественные экосистемы разрушаются, преобразуются, деградируют. Их устойчивость и способность к возобновлению природных ресурсов снижается…
Вмешательство в природные экосистемы.
Любое вмешательство в природу имеет различные последствия, часто непредсказуемые.
Пример: борьба с малярией на о. Северное Борнео (сейчас Бруней). Вот некоторые ее этапы.
1950-е гг. – эпидемия малярии
1955 г. – распыление пестицида диэлдрина для борьбы с комарами . эпидемия прекратилась, комары исчезли, но: погибли другие насекомые, в т.ч. мухи и тараканы погибли маленькие ящерицы, которые жили в домах и питались тараканами погибли кошки, наевшиеся дохлых ящериц быстро размножились крысы - угроза чумы. для исправления ситуации на остров на парашютах сбросили здоровых кошек
Еще примеры «разных времен и народов»:
борьба с кошками как отродьями «диавола» в средневековой Европе и эпидемии чумы, борьба с воробьями в Китае и потери урожая от вредителей, кролики в Австралии, североамериканская норка в СССР и многое др.
«Законы» Коммонера (четыре закона экологии, сформулированные в начале 70-х годов XX в. американским ученым Б. Коммонером в виде афоризмов):
Всё связано со всем.
Всё должно куда-то деваться.
Природа знает лучше — закон имеет двойной смысл — одновременно призыв сблизится с природой и призыв крайне осторожно обращаться с природными системами.
Ничто не даётся даром (вольный перевод — в оригинале что-то вроде «Бесплатных обедов не бывает»).
Естественные экосистемы и принципы построения искусственных экосистем.
Экология рассматривает взаимодействие живых организмов и неживой природы. Это взаимодействие, во-первых, происходит в рамках определенной системы (экологической системы, экосистемы) и, во-вторых, оно не хаотично, а определенным образом организовано, подчинено законам. Экосистемой называют совокупность продуцентов, консументов и детритофагов, взаимодействующих друг с другом и с окружающей их средой посредством обмена веществом, энергией и информацией таким образом, что эта единая система сохраняет устойчивость в течение продолжительного времени.
Экология рассматривает взаимодействие живых организмов и неживой природы. Это взаимодействие, во-первых, происходит в рамках определенной системы (экологической системы, экосистемы) и, во-вторых, оно не хаотично, а определенным образом организовано, подчинено законам. Экосистемой называют совокупность продуцентов, консументов и детритофагов, взаимодействующих друг с другом и с окружающей их средой посредством обмена веществом, энергией и информацией таким образом, что эта единая система сохраняет устойчивость в течение продолжительного времени.
Воздействие человека на окружающую его природную среду может рассматриваться в разных аспектах в зависимости от цели изучения этого вопроса. С точки зрения экологии представляет интерес рассмотрение воздействия человека на экологические системы под углом зрения соответствия или противоречия действий человека объективным законам функционирования природных экосистем. Исходя из взгляда на биосферу как глобальную экосистему, все многообразие видов деятельности человека в биосфере приводит к изменениям: состава биосферы, круговоротов и баланса слагающих ее веществ; энергетического баланса биосферы; биоты. Направленность и степень этих изменений таковы, что самим человеком им дано название экологического кризиса.
Современный экологический кризис характеризуется следующими проявлениями:
- постепенное изменение климата планеты вследствие изменения баланса газов в атмосфере;
- общее и местное (над полюсами, отдельными участками суши) разрушение биосферного озонового экрана;
- загрязнение Мирового океана тяжелыми металлами, сложными органическими соединениями, нефтепродуктами, радиоактивными веществами, насыщение вод углекислым газом;
- разрыв естественных экологических связей между океаном и водами суши в результате строительства плотин на реках, приводящий к изменению твердого стока, нерестовых путей и т.п.;
- загрязнение атмосферы с образованием кислотных осадков, высокотоксичных веществ в результате химических и фотохимических реакций;
- загрязнение вод суши, в том числе речных, служащих для питьевого водоснабжения, высокотоксичными веществами, включая диоксины, тяжелые металлы, фенолы;
- опустынивание планеты;
- деградация почвенного слоя, уменьшение площади плодородных земель, пригодных для сельского хозяйства;
- деградация почвенного слоя, уменьшение площади плодородных земель, пригодных для сельского хозяйства;
- радиоактивное загрязнение отдельных территорий в связи с захоронением радиоактивных отходов, техногенными авариями и т.п.;
- накопление на поверхности суши бытового мусора и промышленных отходов, в особенности практически неразлагающихся пластмасс;
- сокращение площадей тропических и северных лесов, ведущее к дисбалансу газов атмосферы, в том числе сокращению концентрации кислорода в атмосфере планеты;
- загрязнение подземного пространства, включая подземные воды, что делает их непригодными для водоснабжения и угрожает пока еще мало изученной жизни в литосфере;
- массовое и быстрое, лавинообразное исчезновение видов живого вещества;
- ухудшение среды жизни в населенных местах, прежде всего урбанизированных территориях;
- общее истощение и нехватка природных ресурсов для развития человечества;
- изменение размера, энергетической и биогеохимической роли организмов, переформирование пищевых цепей, массовое размножение отдельных видов организмов;
- нарушение иерархии экосистем, увеличение системного однообразия.
Понятие о генетическом паспорте человека.
Понятие о генетическом паспорте человека.
Хотя определить все гены предрасположенности для каждого человека пока невозможно, сегодня уже технически доступен индивидуальный анализ по нескольким десяткам генов. Иными словами, желающие знать свои генетические характеристики могут получить генетический паспорт. В нем должна быть информация о наличии мутаций в генах наследственных болезней и в генах предрасположенности к мультифакторным заболеваниям.
В некоторых западных странах генетическое тестирование проводят по медицинским показаниям или просто в качестве платной услуги. Сведения, записанные в генетическом паспорте, – такая же врачебная тайна, как и результаты любых других медицинских обследований.
Зачем нужен такой паспорт и какие гены необходимо тестировать?
Генетический паспорт обычно делается либо с целью ДНК-идентификации, либо для того, чтобы определить, какие опасные заболевания подстерегают вас в жизни. Это как бы штрих-код, описывающий человека, а точнее наличие тех или иных черт в его ДНК. В США, например, введено генетическое тестирование военнослужащих не для медицинских целей. Его результаты, хранящиеся в специальной базе данных, позволяют идентифицировать человека гораздо точнее, чем надетый на руку жетон. Очень важен генетический паспорт для криминалистики. В той же самой Америке решили пересмотреть дела, связанные с изнасилованием. Сравнили генетические паспорта осужденных и детей, которые родились у потерпевшей. И оказалось, что 30% осужденных по этой статье не являются отцами данных детей.
Если генетический паспорт необходим для здорового молодого человека, желающего знать, какие болезни могут ожидать его в будущем и как предотвратить их развитие, прежде всего нужно составить его родословную и посмотреть, чем болели его родственники.
Носительство мутаций, связанных с моногенными болезнями (фенилкетонурией, муковисцидозом), можно определять, если в семье имеются подобные больные или же заболевание встречается с высокой частотой в той этногеографической группе или того географического региона, где живет обследуемый. Если среди родственников обследуемого есть больше одного человека с одинаковой мультифакторной патологией, необходимо анализировать гены предрасположенности к ее развитию.
При наличии факторов риска, связанных с профессией или особыми условиями среды, нужно тестировать гены, имеющие отношение к болезням, провоцируемым именно этими факторами.
Работы по генетической паспортизации уже начались. Они получили финансовую поддержку в Ирландии, Финляндии, в некоторых штатах США. Большие средства на генетическое тестирование детей с семилетнего возраста выделены правительством Эстонии. Такие работы сопровождаются публикациями в средствах массовой информации, обсуждениями этических и социальных проблем и правовых аспектов. Не случайно в материалах Всемирной организации здравоохранения за 1998 г., посвященных этическим аспектам медицинской генетики, подчеркивается, что пресимптоматическое тестирование можно проводить только по желанию, только в случае возможной реальной пользы для пациента или его родственников, при условии максимально объективного информирования пациента о результатах тестирования.
Большинство людей (около 70%) уже теперь хотели бы получить генетический паспорт, 20% не хотят этого, 14% затруднились ответить. Большинство хотели бы получить медико-генетическую информацию о себе (60%) и своем супруге (50%). 92% респондентов считают, что такую информацию должны иметь родители о несовершеннолетних детях; 87% – взрослые дети о родителях; 50% считают, что о генетических рисках здоровья должен знать супруг; 73% доверились бы врачам медицинских учреждений при соблюдении врачебной тайны. Более 80% опрошенных –против предоставления этой информации руководству предприятий.
Билет № 9
Современные методы исследования генома.
Современные методы секвенирования ДНК (модификации метода Сэнгера для автоматического секвенирования, пиросеквенирование).
Секвенирование биополимеров (белков и нуклеиновых кислот – ДНК и РНК) – это определение их первичной аминокислотной или нуклеотидной последовательности. Для секвенирования применяются методы Эдмана, Сэнгера, Максама– Гилберта и др.
С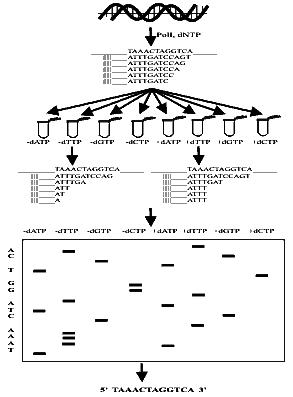 еквенирование
ДНК по Сэнгеру: "плюс-минус" метод
(рисунок 1).
В
качестве матрицы в реакции полимеразного
копирования использовался одноцепочечный
фрагмент ДНК, в качестве праймеров –
синтетические олигонуклеотиды или
природные субфрагменты, получаемые при
гидролизе рестрицирующими эндонуклеазами,
а в качестве фермента – фрагмент Кленова
ДНК полимеразы I (PolI) из E.coli.
Метод включает два этапа. Сначала в
ограниченных условиях проводят
полимеразную
реакцию
в присутствии всех четырех типов dNTP
(один из них мечен по альфа-положению
фосфата), получая на выходе набор
продуктов неполного копирования
матричного фрагмента. Смесь очищают от
несвязавшихся дезоксинуклеозидтрифосфатов
и делят на восемь частей. После этого в
"плюс"-системе проводят четыре
реакции в присутствии каждого из четырех
типов нуклеотидов, а в "минус"-системе
– в отсутствие каждого из них. В
результате, в "минус"- системе
терминация
происходит перед dNTP данного типа, а в
"плюс"-системе – после него.
Полученные таким образом восемь образцов
разделяют с помощью электрофореза,
"считывают" сигнал и определяют
последовательность исходной ДНК. Этим
способом была секвенирована короткая
ДНК фага фХ174, состоящая из 5386 нуклеотидных
пар.
еквенирование
ДНК по Сэнгеру: "плюс-минус" метод
(рисунок 1).
В
качестве матрицы в реакции полимеразного
копирования использовался одноцепочечный
фрагмент ДНК, в качестве праймеров –
синтетические олигонуклеотиды или
природные субфрагменты, получаемые при
гидролизе рестрицирующими эндонуклеазами,
а в качестве фермента – фрагмент Кленова
ДНК полимеразы I (PolI) из E.coli.
Метод включает два этапа. Сначала в
ограниченных условиях проводят
полимеразную
реакцию
в присутствии всех четырех типов dNTP
(один из них мечен по альфа-положению
фосфата), получая на выходе набор
продуктов неполного копирования
матричного фрагмента. Смесь очищают от
несвязавшихся дезоксинуклеозидтрифосфатов
и делят на восемь частей. После этого в
"плюс"-системе проводят четыре
реакции в присутствии каждого из четырех
типов нуклеотидов, а в "минус"-системе
– в отсутствие каждого из них. В
результате, в "минус"- системе
терминация
происходит перед dNTP данного типа, а в
"плюс"-системе – после него.
Полученные таким образом восемь образцов
разделяют с помощью электрофореза,
"считывают" сигнал и определяют
последовательность исходной ДНК. Этим
способом была секвенирована короткая
ДНК фага фХ174, состоящая из 5386 нуклеотидных
пар.
Секвенирование ДНК по Сэнгеру: метод "терминаторов". Как наиболее мощный и технологичный, этот способ, несколько модифицированный, применяется до сих пор. В основе метода тоже лежит ферментативное копирование с помощью фрагмента Кленова ДНК полимеразы I из E.coli (рисунок 2). В качестве праймеров используются синтетические олигонуклеотиды. Специфическую терминацию синтеза обеспечивают добавлением в реакционную смесь помимо четырех типов dNTP (один из которых радиоактивно мечен по альфа положению фосфата) еще и одного из 2',3'-дидезоксинуклеозидтрифосфатов (ddATP, ddTTP, ddCTP или ddGTP), способного включаться в растущую цепь ДНК, но не способного обеспечивать дальнейшее копирование из-за отсутствия 3'-ОН группы. Соотношение концентраций dNTP/ddNTP авторы подбирают экспериментально так, чтобы в итоге получить набор копий ДНК различной длины. Таким образом, для определения первичной структуры исследуемого фрагмента ДНК требуется провести четыре реакции копирования: по одному типу терминаторов в каждой реакции. После этого полученные продукты разгоняются в полиакриламидном геле на соседних дорожках и по расположению полос определяется последовательность нуклеотидов.
Секвенирование ДНК по Максаму–Гилберту: метод химической деградации. Метод основан на специфической химической деградации фрагмента ДНК, радиоактивно меченного с одного конца. Препарат меченной ДНК разделяют на четыре части (или аликвоты) и каждую обрабатывают реагентом, модифицирующим одно или два из четырех оснований. А. Максам и У. Гилберт предложили модифицировать пуриновые основания диметилсульфатом. При этом происходит метилирование адениновых остатков по азоту в положении 3, а гуаниновых – по азоту – в положении 7. Обработка образца ДНК соляной кислотой при 0°С приводит к выщеплению метиладенина. Последующая инкубация при температуре 90°С в щелочной среде вызывает разрыв сахарно-фосфатной цепи ДНК в местах выщепления оснований. Обработка пиперидином приводит к гидролизу образца по остаткам метилгуанина. Пиримидиновые основания модифицируют гидразином. Если реакцию вести в бессолевой среде, то модифицируются как цитозин, так и тимидин; если обработку вести в присутствии 2М NaCl, то модифицируется лишь цитозин. Расщепление цепи ДНК на фрагменты и в этом случае осуществляется пиперидином. Условия реакций авторы подбирали таким образом, чтобы в итоге получить полный набор субфрагментов разной длины. Последующий электрофорез в полиакриламидном геле позволяет восстановить полную структуру исследуемого фрагмента.
Автоматическое секвенирование ДНК. В основу автоматического секвенирования заложен уже упоминавшийся метод ферментативного секвенирования с использованием терминирующих ddNTP (*). Как и классический вариант Сэнгера, автоматическое секвенирование включает две стадии: проведение терминирующих реакций и разделение их продуктов с помощью электрофореза. Как правило, автоматизирована лишь вторая стадия, т.е. разделение меченных фрагментов ДНК в ПААГ, получение спектра эмиссии флуорофоров и последующая обработка собранных данных. Таким образом, автоматическое секвенирование идеологически отличается от современного ему ручного секвенирования только типом используемой метки.
Cеквенаторы. Современные секвенаторы можно разделить по типу проводимого электрофореза на капиллярные и те, в которых разделение происходит в гелевых пластинах. Последние могут также различаться по количеству детектируемых красителей: один, два или четыре. Капиллярные секвенаторы выпускаются только в варианте, использующем детекцию четырех флуоресцентных красок.
Наиболее успешным секвенатором начала 1990-х гг. стал ABI 373 (Applied Biosystems). В этой модели используется одновременная детекция четырех красителей (TAMRA, FAM, ROX, JOE), возбуждаемых излучением аргонового лазера. Среди секвенаторов, детектирующих флуоресценцию одного красителя, можно выделить секвенатор MicroGene Blaster фирмы Visible Genetics, который позволяет одновременно проводить анализ четырех образцов, меченных Cy5.5, на 16 дорожках.
Секвенаторы, работающие на двух красителях в инфракрасной области спектра, выпускает фирма LI-COR. Ее основная машина IR2 DNA Sequencer. Конкурентом IR2 DNA Sequencer стал секвенатор ABI Prism 377, который пришел на смену ABI 373. Основное новшество этой машины заключается в замене оптической фильтрации флуоресценции, которая использовалась в ABI 373 для разделения спектров испускания красителей, на "виртуальную" фильтрацию. Схожая технология записи и обработки данных используется в другом секвенаторе фирмы Applied Biosystems – ABI Prism 310. Это однокапиллярный секвенатор, способный в автоматическом режиме последовательно просканировать 96 образцов.
ABI Prism 310 стал пионером капиллярной технологии в секвенировании. Нынешнее поколение капиллярных секвенаторов представляет собой уже мультикапиллярные машины, некоторые из которых демонстрируют гораздо более высокую производительность даже в сравнении с ABI 377. В зависимости от поставленных задач можно остановить свой выбор на восьмикапиллярном Beckman CEQ 2000 (Beckman Coulter), шестнадцатикапиллярном секвенаторе ABI Prism 3100 или 96-капиллярных ABI Prism 3700 и MegaBACE 1000 (Amersham-Pharmacia-Biotech-Molecular Dynamics).
Человек как фактор эволюции Homo sapiens.
Генная реконструкция филогенеза человека

Билет № 10
Промышленная биотехнология. Основные методы культивирования микроорганизмов. Получение кормовых белковых продуктов, медицинских, ферментных препаратов и метаболитов бактерий первой и второй фазы роста.
Промышленная микробиология – это наука о получении различных целевых продуктов на основе жизнедеятельности микроорганизмов. Промышленная микробиология (или техническая микробиология) в настоящее время представляет собой также самостоятельную и наиболее крупнотоннажную отрасль современной промышленной биотехнологии. Огромное разнообразие микроорганизмов, утилизирующих в качестве ростовых субстратов различные соединения, в том числе отходы, позволяет получать широкий спектр биологически активных соединений, а также осуществлять полезные для человека реакции, включая обезвреживание отходов, трансформацию и получение энергии, и многое другое.
В настоящее время в различных процессах промышленной микробиологии получают около 200 соединений, обладающих коммерческой ценностью. Важнейшими среди них являются: алкалоиды, аминокислоты, антибиотики, антиметаболиты, антиоксиданты, белки, витамины, гербициды, инсектициды, коферменты, липиды, нуклеиновые кислоты, органические кислоты, пигменты, ПАВ, полисахариды, полиоксиалканоаты, противоопухолевые агенты, растворители, сахара, стерины, ферменты, нуклеотиды, нуклеозиды, эмульгаторы.
Принципиальное отличие биотехнологических процессов от чисто химических заключается в следующем:
чувствительность биологических агентов к физико-механическим воздействиям;
наличие межфазового переноса веществ (по типу «жидкость – клетки», «газ – жидкость – клетки»);
требования условий асептики;
низкие скорости протекания многих процессов в целом;
нестабильность целевых продуктов;
пенообразование;
сложность механизмов регуляции роста и биосинтеза.
Ф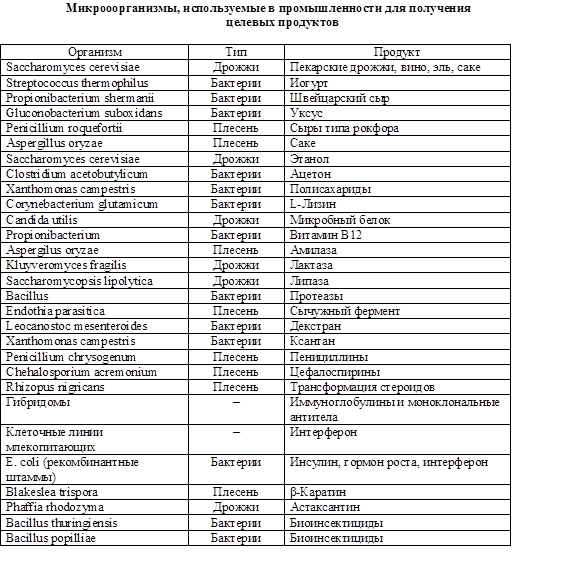 акторами,
лимитирующими рост микробной популяции,
являются нехватка питательных веществ
и отравление продуктами собственной
жизнедеятельности. Если постоянно
обновлять в культиваторе среду, подводя
свежий раствор питательных веществ и
удаляя отработанную культуральную
жидкость вместе с избытком биомассы,
можно сделать популяцию практически
«бессмертной», способной размножаться
в течение любого необходимого исследователю
времени. Идея и разработка метода
непрерывного культивирования
микроорганизмов принадлежат французскому
микробиологу Ж. Моно и американским
ученым А. Новику и Л. Сциларду.
акторами,
лимитирующими рост микробной популяции,
являются нехватка питательных веществ
и отравление продуктами собственной
жизнедеятельности. Если постоянно
обновлять в культиваторе среду, подводя
свежий раствор питательных веществ и
удаляя отработанную культуральную
жидкость вместе с избытком биомассы,
можно сделать популяцию практически
«бессмертной», способной размножаться
в течение любого необходимого исследователю
времени. Идея и разработка метода
непрерывного культивирования
микроорганизмов принадлежат французскому
микробиологу Ж. Моно и американским
ученым А. Новику и Л. Сциларду.
В бывшем Советском Союзе пионером этого наиболее «технологичного» метода биологических исследований явился Николай Дмитриевич Иерусалимский, выдающийся советский микробиолог, академик АН СССР, организатор Института биохимии и физиологии микроорганизмов АН СССР в Пущине.
Математические аспекты автоселекции при непрерывном культивировании микробов с середины 1960-х гг. интенсивно разрабатывались в Красноярске членом-корреспондентом АН СССР Иваном Александровичем Терсковым в Институте физики им. Л.В.Киренского СО РАН, а затем его учеником д.б.н. Н.С. Печуркиным в Институте биофизики СО РАН.
Управление ростом микробной популяции может осуществляться при непрерывном, или проточном, культивировании различными способами:
в режиме хемостата управляющим параметром служит концентрация питательных веществ, лимитирующая неограниченное размножение микроорганизмов;
в режиме без внешнего лимитирования работает, например, турбидостат, где постоянной задается плотность биомассы, или
рН-стат, где постоянна концентрация водородных ионов.
Ферментеры
Ферментеры с подводом энергии к газовой фазе (группа ФГ). Их общий признак – подвод энергии в аппарат через газовую фазу, которая является ее носителем.
Ферментеры с вводом энергии жидкой фазой (группа ФЖ) наиболее сложны по конструкции и энергоемки, но обеспечивают наиболее вы-сокие по сравнению с группой ферментеров ГФ значения коэффициента массопередачи кислорода, свыше 6 кг/м3 ч.
Ферментеры с подводом энергии газовой и жидкой фазами (группа ФЖГ). Основными их конструкционными элементами являются перемешивающие устройства всех известных типов, а также наличие в совокупности насосов и перемешивающих устройств.
Получение кормовых белковых продуктов
Наиболее дефицитным компонентом пищи является белок, в особенности, – высокой биологической ценности, то есть животного происхождения. Мировая потребность в белка в настоящее время удовлетворяется примерно на 40 %. Предполагалось, что к 2000 г. с ростом населения потребность в белке увеличится, при этом дефицит кормового белка возрастет до 147 %. Поэтому изыскание эффективных способов увеличения ресурсов белка для прямого или непрямого (через организм сельскохозяйственных животных) увеличения пищевых ресурсов является одной из основных задач научно-технического прогресса.
Нетрадиционным и принципиально новым способом получения белковых веществ является микробиологический синтез. По скорости роста микроорганизмы превосходят сельскохозяйственные культуры в сотни, а животных – в тысячи раз. Поэтому микробиологический синтез с большей эффективностью использует материальные и энергетические ресурсы, не требует больших земельных площадей и не зависит от погодных и климатических условий и не загрязняет окружающую среду ядохимикатами, так как не использует пестициды. Качество микробных белков близко белкам животного происхождения. Применение микробных белков в кормопроизводстве улучшает качество и усвояемость традиционных растительных кормов. Например, 1 т кормовых дрожжей обеспечивает экономию 5 т зерна и увеличивает продуктивность в животноводстве на 15–30 %.
Современный средний завод по производству микробного белка мощностью 50 т/год и занимающий 0,2 га может обеспечить потребность в белке до 10 млн. человек. Сельскохозяйственные технологии для таких масштабов производства требуют до 16 тыс. га, засеянных пшеницей, либо содержание фермы с производительностью 400 поросят/день.
В 1960-е гг. появился новый термин – «белок одноклеточных» (single cell protein, «SCP»), означающий целые неживые высушенные микробные клетки (водорослей, дрожжей, бактерий, грибов), предназначенные в качестве белкового продукта для кормовых и пищевых целей. А саму идею использования биомассы микроорганизмов в качестве белковых компонентов питания с 1890 г. начал пропагандировать Дельбрюк, который вместе с коллегами разработал первый технологический процесс выращивания пивных дрожжей Saccharomyces cerevisiae на мелассе.
В настоящее время в микробиологических производствах белка применяется различное сахаросодержащее сырье. Это отходы пищевой, молочной, спиртовой, сахарной и целлюлозной промышленности и продукты переработки растительного сырья (древесины, соломы, торфа, несъедобных частей растений – стебли, лузга, кочерыжки). Питательные среды, приготовленные на основе перечисленных субстратов, содержат наборы моно- и дисахаров, органические кислоты, спирты и другие органические соединения, а также минеральные элементы, то есть являются сложными многокомпонентыми субстратами. Поэтому при их применении используют штаммы-продуценты, способные, во-первых, усваивать как пентозы, так и гексозы, и, во-вторых, – устойчивые к присутствию спиртов, фурфурола и других продуктов гидролиза растительных биомасс. Наибольшее распространение получили виды дрожжей рода Candida: C. utilis, C. scottii, C. tropicalis, способные утилизировать наряду с гексозами пентозы и толерантные к наличию фурфурола в среде.
Эффективным белковым продуктом оказались цианобактерии рода Spirulina, растущие в природных условиях и способные фиксировать атмосферный азот. Биомасса Spirulina содержит (%): до 70 белков, полноценного аминокислотного состава, 19 углеводов, 4 нуклеиновых кислот и 4 липидов, 6 пигментов и по 3 золы и волокон. Клеточная стенка имеет отличный от микроводорослей состав и легко переваривается. Низкий уровень нуклеиновых кислот в биомассе, нетоксичность пигментов фикоцианинов, высокий уровень переваримого белка, – все это сделали данную биомассу полноценным белковым продуктом пищевого назначения.
В середине 1970-х гг. активизировались исследования, направленные на разработку технологий получения микробного белка с использованием хемолитоавтотрофных микроорганизмов. Хемолитоавтотрофные водородокисляющие бактерии используют в качестве источника углерода углекислоту, а энергии – реакцию окисления водорода. Перспективность водородокисляющих бактерий определяется их автотрофией и независимостью от дефицитных источников органического сырья, быстрым ростом, высоким содержанием полноценного по аминокислотному составу белка, отсутствием внеклеточных промежуточных продуктов обмена органической природы (единственным побочным продуктом процесса окисления водорода является вода), высокой экологической чистотой процесса производства и получаемого продукта.
Получение бактериальных удобрений и метаболитов бактерий первой и второй фазы роста
Микроорганизмы играют большую роль в повышении плодородия почвы, так как в процессе роста и развития улучшают ее структуру, обогащают питательными веществами, способствуют более полному использованию удобрений. Интенсивное растениеводство обедняет почву азотом, так как значительная его доля ежегодно выносится из почвы вместе с урожаем. С древних времен для восстановления и улучшения почв существует практика ииспользования бобовых растений, способных в симбиозе с азотфиксирующими микроорганизмами восполнять почвенные запасы азота в результате диазотрофности (усвоения атмосферного азота). Большой положительный эффект от возделывания бобовых вызвал постановку исследований явления диазотрофности.
Практическое применение нашли также симбиотические бактерии рода Rhizobium, развивающиеся в клубеньках бобовых растений. Как только была выяснена роль симбиотических бактерий рода Rhizobium в азотфиксации, стали разрабатывать способы внесения этих микроорганизмов в почву и также для инокуляции семян. Затраты на применение этих способов невелики, техника применения весьма проста, а эффект от их применения значителен. Культивирование бобовых, положительно влияя на азотный баланс почв, также облегчает борьбу с эрозией и помогает восстанавливать истощенные земли.
Клубеньковые бактерии обладают избирательной способностью по отношению к растению-хозяину. Эта особенность азотфиксаторов положена в основу их классификации внутри рода Rhizobium. Так, для бактерий Rh. leguminosarum растением-хозяином являются горох, вика, кормовые бобы, чина, чечевица; для Rh. phaseoli – фасоль; Rh. japonicum – соя; Rh. trifolii – клевер; Rh. vigna – вигна, маис, арахис и др. Процесс азотфиксации протекает только в клубеньках на корнях бобовых растений, которые образуются в результате проникновения бактерий через корневые волоски в корень.
Первая коммерческая разновидность культуры для инокуляции семян (товарное название «Nitragin») была запатентована в Великобритании Ноббе и Хилтнером (1896). Для разных бобовых в то время выпускали 17 вариантов культуры. Бактерии выращивали на агаризованных средах, далее соскабливали с поверхности плотной среды и суспендировали в молоке. Суспензию бактерий выливали на кучу семян, перемешивали и далее семена высушивали в тени. Вскоре семена высевали. Данный метод пригоден для инокуляции сравнительно небольших объемов семян и применялся во многих странах с конца тридцатых до начала семидесятых годов XX в.
В настоящее время для поддержания жизнеспособности симбиотических азотфиксирующих бактерий используют разнообразные носители, но лучшим считается торф. Сухие препараты азотфиксаторов, приготовленные на основе клубеньковых бактерий рода Rhizobium и предназначенные для повышения урожайности бобовых растений (гороха, фасоли, сои, клевера, люцерны, люпина и др.) в настоящее время выпускаются под товарным названием «Нитрагин».
Аналогом азотных удобрений является другой препарат азотфиксирующих бактерий – «Азотобактерин», который выпускается промышленностью в нескольких вариантах. Бактерии рода Azotobacter являются свободноживущими азотфиксирующими микроорганизмами и обладают высокой продуктивностью азотфиксации (до 20 мг/г использованного сахара). Помимо связывания атмосферного азота, эти бактерии продуцируют биологически активные соединения (витамины, гиббериллин, гетероауксин и др.). В результате этого инокуляция азотобактерином стимулирует прорастание семян и ускоряет рост и развитие растений. Более того, Azotabacter способен экскретировать фунгицидные вещества. Этим угнетается развитие в ризосфере растений микроскопических грибов, многие из которых тормозят развитие растений. Технология получения сухого препарата азотобактерина аналогична получению сухого нитрагина и включает получение посевного материала и культивирование бактерий в контролируемых условиях в глубинной стерильной культуре до начала стационарной фазы. Готовый препарат с содержанием не менее 5 млрд. жизнеспособных клеток на 1 кг при остаточной влажности 5–7 % фасуют в полиэтиленовые мешки 0.4–2.0 кг, которые герметизируют и далее хранят при температуре до 15°С. Промышленностью выпускаются также торфяной и почвенный препараты азотобактерина
В последнее десятилетие XX в. для изучения биологической азотфиксации стали применять методы молекулярной биологии и новейшие методы генетики. Установлена возможность с помощью колифага P1 размножать свободноживущую азотфиксирую-щую бактерию Klebsiella pneumoniae М5 и с ее помощью трансдуцировать nif-гены (гены азотфиксации). Также доказано, что перенос nif-генов возможен с помощью плазмид от штамма-азотфиксатора к штамму, не обладающему диазотрофностью. Обнаружены конъюгативные плазмиды, несущие гены азотфиксации, относительно легко передающиеся при конъюгации от штамма к штамму.
Использование отдельных групп микроорганизмов в пищевой промышленности
Спектр продуктов питания, получаемых при помощи микроорганизмов, обширен. Это продукты, получаемые в результате брожения - хлеб, сыр, вино, пиво, творог и так далее. До недавнего времени биотехнология использовалась в пищевой промышленности с целью усовершенствования освоенных процессов и более умелого использования микроорганизмов, но будущее здесь принадлежит генетическим исследованиям по созданию более продуктивных штаммов для конкретных нужд, внедрению новых методов в технологии брожения.
Мы рассмотрим использование отдельных групп микроорганизмов: при получении молочнокислых продуктов, в хлебопечении, в мясной и рыбной промышленности.
Агроэкосистемы как яркий пример антропогенных экосистем.
Сельскохозяйственные системы – экосистемы на ранних стадиях сукцессий (высокопродуктивные, но сильно упрощенные). Проблема – вторжение других видов (сорняков, вредителей, болезней). Требуются большие затраты энергии и ресурсов для поддержания таких экосистем.
Задача – сохранить баланс между упрощенными антропогенными экосистемами и соседствующими более сложными природными экосистемами, от которых в итоге зависят антропогенные экосистемы.
Сельскохозяйственные районы весьма различны по природным условиям, типам землепользования и степени освоения. Тем не менее экологические проблемы в них имеют много общего. Это связано со следующими обстоятельствами:
охватом антропогенными нагрузками больших площадей, иногда практически на 100%;
малой лесистостью и небольшими площадями лугово-степных участков;
значительной обнаженностью, дефдированностью и эродированностью почвенного покрова;
преобладанием определенных видов загрязнения в почве, воде и грунтах, связанных с удобрениями.
Перечисленные обстоятельства свиде-тельствуют о специфике экологического состояния сельскохозяйственных райо-нов, о правомерности выделения “агроэкологического” типа оценок территории.
Основной аспект агроэкологической оценки - анализ условий развития сельскохозяйственных растений: их роста, фенологии, урожайности, отно-шения к удобрениям, болезням, сезонным изменениям условий тепла и влаги - морозам, заморозкам, засухам, переувлажнению.
Экологические условия сельскохозяйственных угодий наиболее изменчивы на площадях богарного, неполивного земледелия. Более стабильны они в зонах орошения, где мероприятия по мелиорации ослабляют влияние внешних условий.
При региональной оценке районов сельского хозяйства важно определить степень устойчивости экосистем к антропогенным нагрузкам. Устойчивость повышается от песчаных грунтов к глинистым, от щелочных почв к кислым, при снижении континентальности климата, нарастании годового увлажнения и увеличении биологической продуктивности фитоценозов - как естественных, так и культурных.
Большая устойчивость угодий западных и северо-западных районов России к антропогенным нагрузкам не всегда имеет решающее значение для экологического состояния. Дело в том, что этим районам характерны более интенсивные типы землепользования, большие дозы вносимых удобрений. Максимальная интенсификация хозяйства характерна для территорий, прилегающих к крупным городам и промышленным зонам (Москва, Санкт-Петербург), которых также больше в западных районах. Очевидно, объективная оценка экологического состояния возможна лишь при равном учете природных и экономических факторов.
Кардинальные изменения природной среды сельскохозяйственных районов обусловлены тем, что на площадях угодий меняются потоки вещества, нарушается твердый, жидкий и растворенный сток. Сведение лесов увеличивает смыв почвы, твердый сток рек, приводит к заилению русел, водохранилищ, пойменных массивов. Расходы водотоков при сокращении лесных площадей на 10% снижаются в среднем на 5%. Активная миграция элементов по склонам, их быстрое поступление в водоемы с одновременным сокращением стока приводит к сильному загрязнению поверхностных вод. Это загрязнение может быть токсичным, поскольку такие опасные элементы, как кадмий, ртуть, стронций, свинец, цинк, относятся к наиболее подвижным в большинстве видов почв.
Прилегающие к крупным населенным пунктам сельскохозяйственные районы на площадях в сотни кв. км испытывают на себе влияние промышленного загрязнения. Наибольшую роль здесь играет загрязнение серой, которая в виде сернистых соединений легко разносится воздушными потоками. В нормально увлажненных нейтральных почвах влияние этого вида загрязнения невелико, но в кислых оно усиливает подкисление. На переувлажненных почвах, особенно на поймах, это может привести к резкому закислению после осушения.
Основные изменения почв в земледелии связаны с механическим воздействием на нее и с внесением удобрений. Вспашка меняет профиль почвы, разрушает структуру, приводит к обеднению верхних горизонтов, способствует усилению водной эрозии и дефляции. Наряду с рыхлением идет и уплотнение почвы.
Велико также значение органических и минеральных удобрений, мировое потребление которых - около 90 млн. т в год. Удобрения не только компенсируют вынос из почвы азота, фосфора и калия, но нередко оказываются избыточными, заражают подземные и поверхностные воды. Это имеет место главным образом в развитых странах, где вносится более 100 кг/га. В развивающихся странах этот показатель в 5 раз ниже.
Получение высоких урожаев в настоящее время невозможно без использования различных ядохимикатов для защиты растений - пестицидов, потребление которых превышает 4 млн. т/год. Однако сейчас их использование сокращается в связи с приспособлением к ним многих вредителей, гибелью почвенных микроорганизмов, заражением овощных культур и накоплением ядовитых веществ в поверхностных водах, донных осадках водоемов, организмах животных и человека.
Чрезмерные антропогенные нагрузки приводят к напряженной экологической обстановке во многих районах сельскохозяйственного освоения.
Сравнительная характеристика природных экосистем и агроэкосистем.
Понятие агроэкосистемы воспринимается неоднозначно. К примеру, по мнению Ю. Одума, агроэкосистемы — это одомашненные экосистемы, которые во многих отношениях занимают промежуточное положение между природными экосистемами (луга, леса) и искусственными (города). Другой американский агроэколог Р. Митчелл считает, что подобно тому, как морские свинки — это не обитатели моря и не представители отряда парнокопытных, так и агроэкосистемы — это не настоящие экосистемы, но и не самодовлеющие сельскохозяйственные единицы. Во всех агроэкосистемах экономические соображения влияют на структуру посевов и набор культур.
Некоторые исследователи считают, что роль человека, под управлением которого находится агроэкосистема, настолько значительна, что следует говорить об артеприродной основе агроэкосистем. Действительно, агроэкосистемы сходны с урбанизированными и промышленными системами своей зависимостью от внешних факторов, т. е. от окружающей среды на входе и выходе системы. Однако в отличие от них агроэкосистемы по преимуществу автотрофны.
В свете современных представлений агроэкосистемы (агробиогеоценозы) — вторичные, измененные человеком биогеоценозы, ставшие значительными элементарными единицами биосферы; их основу составляют искусственно созданные, как правило, обедненные видами живых организмов биотические сообщества. Эти сообщества формируют и регулируют люди для получения сельскохозяйственной продукции. Агроэкосистемы отличаются высокой биологической продуктивностью за счет привнесения дополнительной энергии и доминированием одного или нескольких избранных видов (сортов, пород) растений или животных. Выращиваемые культуры и разводимые животные подвергаются искусственному, а не естественному отбору. Как экологические системы агроэкосистемы неустойчивы: у них слабо выражена способность к саморегулированию, без поддержки человеком они быстро распадаются или дичают и трансформируются в естественные биогеоценозы (например, мелиорированные земли — в болота, насаждения лесных культур — в лес).
Агроэкосистемы с преобладанием зерновых культур существуют не более одного года, многолетних трав — 3–4 года, плодовых культур — 20–30 лет, а затем они распадаются и отмирают. Лесные полезащитные полосы, являющиеся элементами агроэкосистем, в степной зоне существуют не менее 30 лет. Однако без поддержки человеком (рубки ухода, дополнения) они постепенно «дичают», превращаясь в естественные экосистемы, или погибают. Преобладающая разновидность агроэкосистем — искусственные фитоценозы: окультуренные (планомерно эксплуатируемые луга и пастбища); полукультурные (непостоянно регулируемые искусственные насаждения — сеяные, многолетние луга); культурные (постоянно регулируемые многолетние насаждения, полевые и огородные культуры); интенсивно культурные (парниковые и оранжерейные культуры, гидропоника, аэропоника и другие, требующие создания и поддержания особых почвенных, водных и воздушных условий). Управление агроэкосистемой осуществляется извне и подчинено внешним целям.
Заслуживает внимания определение Р.А. Полуэктова, назвавшего агроэкосистемы специальным видом экосистем сельскохозяйственного поля, где произрастают культурные растения, обитают другие виды растений и животных и происходит сложная цепь физических и химических трансформаций энергии и вещества.
Билет № 11
Явление РНК-интерференции, его фундаментальные и прикладные аспекты.
Нобелевская премия в области медицины и физиологии за 2006 г. присуждена американским генетикам: 47-летнему профессору патологии и генетики Медицинской школы Стэнфордского университета Эндрю Файеру (Andrew Z. Fire) и 45-летнему профессору молекулярной медицины Медицинской школы Массачусетского университета в Вустере Крэйгу Меллоу (Craig C. Mello). Исследователи открыли и описали механизм универсального явления – «генной цензуры», или РНК-интерференции, при которой происходит избирательное подавление экспрессии генов, унаследованных организмом. Это один из редких случаев, когда от начала работ, до их увенчания Нобелевской премией прошло чуть больше десяти лет.
Первые загадочные результаты были получены еще в 1980-х – начале 1990-х гг. Сначала в опытах на кишечной палочке было установлено, что искусственное введение в клетку синтетических фрагментов одноцепочечных молекул рибонуклеиновой кислоты может приводить к отключению генов. Затем на цветах (петуния) и червях (нематода Caenorhabditis elegans) введение в клетки дополнительных копий генов вместо усиления экспрессии наблюдался противоположный эффект: полное "замолкание".
В 1997 г. Э.Файер и К.Меллоу в экспериментах с нематодой Caenorhabditis elegans обнаружили, что РНК очень эффективно отключает гены, если ее вводить короткими фрагментами, причем не одноцепочечными, а двухцепочечными! Это наблюдение было сделано почти случайно – исследователи вовсе не предполагали, что такие фрагменты будут работать лучше одноцепочечных, они вводили их просто для контроля. Поначалу это явление казалось загадочным, т.к. двухцепочечная РНК сама по себе не может связаться с матричной, ей сначала надо расплестись на две нити. Первооткрыватели тоже не могли его объяснить, это удалось сделать лишь позже. Впервые об открытии было объявлено в феврале 1998 г., когда Э.Файер и К.Меллоу опубликовали в журнале «Nature» статью о новом явлении. Эффект "гашения" экспрессии определенных генов малыми РНК был назван К.Меллоу РНК-интерференцией, а молекулы, вызывающие его, назвали siRNA (small interfering ribonucleic acids – малые интерферирующие рибонуклеиновые кислоты).
Структурно-функциональная организация микро РНК, коротких интерферирующих Р НК и других малых РНК. Биогенез малых РНК.
В класс малых РНК включают молекулы, содержащие от 20 до 300 нуклеотидов. За эффект РНК-интерференции отвечают самые короткие из них – siRNA (Малые интерферирующие РНК), состоящие всего из 21–28 (у млекопитающих из 21–23) нуклеотидов (таблица 3). Особенность этих молекул проявляется в том, что они в отличие от большинства других клеточных РНК, состоящих всего из одной цепи нуклеотидов, являются двунитчатыми. Нуклеотиды с противоположных нитей (цепей) siRNA спариваются по тем же законам комплементарности, которые формируют двунитчатые цепи ДНК в хромосомах. Кроме того, по краям каждой из цепей siRNA всегда остаются два неспаренных нуклеотида.
Происхождение двухцепочечных РНК в клетке может быть различным: транскрипция с конаправленных промоторов, активность клеточной или вирусной РНК-зависимой РНК-полимеразы, самокомплементарные транскрипты, способные к внутримолекулярному спариванию и образованию шпилечных структур, а также искусственно введенные в клетку молекулы.
Механизм РНК-интерференции очень древний. Скорее всего, биологическая эволюция создала его более миллиарда лет назад, когда нашу планету населяли только микроорганизмы. С его помощью они получили возможность защищаться от нападений вирусов. Позже эту способность переняли у них грибки и растения, а потом и животные. Многоклеточные организмы также научились прицельно отключать с помощью РНК-интерференции собственные гены, прерывая процесс синтеза кодируемых ими белков.
Класс |
Подкласс |
Длина (в нуклеотидах) |
Биогенез |
Механизм действия |
Биологическая функция |
Микро РНК (miRNA) |
Нет |
~22(19–25) |
Двухстадийное разрезание предшественников шпилечных структур ферментами Drosha и Dicer |
Репрессия транс-ляции, разрезание мРНК |
Различные (часто дейст-вуют в процессе развития и клеточной дифференцировки) |
Короткая интерферирую-щая РНК (siRNAs) |
Эндогенная взаимодей-ствующая siRNA |
21–22 (у не-матоды) 21 (у расте-ний) |
Разрезание длинных эндогенных dsRNAs с помощью Dicer |
Разрезание мРНК |
Неизвестна |
Ассоцииро-ванная с повторами siRNA (rasiRNA) |
24–24 (у растений) 24–27 (у плодовой мушки) |
Разрезание длинных dsRNAs, происходящих от повторяющихся последовательностей, ферментом Dicer |
Модификация гистона и/или ДНК |
Сайленсинг транспозонов, повторяющихся генов и вирусов |
|
Малая ска-нирующая РНК (scnRNA) |
~28 |
Разрезание длинных dsRNAs с помощью Dicer |
Метилирование гистонов, ведущее к элиминации ДНК |
Перестройка генома в ходе конъюгации |
|
Крошечная некодирующая РНК (tncRNA) |
Нет |
~20 |
Производится ферме-том Dicer из неизвестного предшественника |
Неизвестен |
Неизвестна |
Малая моду-ляторная РНК (smRNA) |
Нет |
~20 |
Неизвестен |
Транскрипционная трансактивация |
Дифференциация нейронов |
Н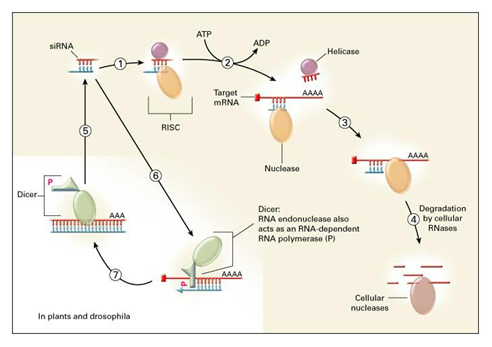 а
рисунке 6 схематически показан механизм
работы siRNA
в клетках растений и животных. Первые
4 этапа являются общими для всех
организмов. Сначала с молекулой siRNA
связываются белки-ферменты хеликаза
и нуклеаза (см. этап 1), формируя комплекс
RISC
(RNA-induced
silencing
complex;
silence
– англ. молчать, замолкать; silencing
– замолкание, так в англоязычной и
специальной литературе называют процесс
"выключения" гена).
а
рисунке 6 схематически показан механизм
работы siRNA
в клетках растений и животных. Первые
4 этапа являются общими для всех
организмов. Сначала с молекулой siRNA
связываются белки-ферменты хеликаза
и нуклеаза (см. этап 1), формируя комплекс
RISC
(RNA-induced
silencing
complex;
silence
– англ. молчать, замолкать; silencing
– замолкание, так в англоязычной и
специальной литературе называют процесс
"выключения" гена).
Хеликаза раскручивает нити siRNA, в результате чего они расходятся (этап 2). Одна из нитей, к которой прикреплен фермент нуклеаза, – ее официальное название аргонавт (argonaute), может теперь связаться с комплементарным (строго соответствующим ей) участком однонитчатой мРНК (этап 3), позволяя нуклеазе разрезать ее. Разрезанные же участки мРНК подвергаются действию других клеточных РНКаз, которые разрезают их на более мелкие куски (этап 4).
У растений и насекомых выработалась своеобразная система усиления защитного действия siRNA (этапы 5–7). Присоединяясь к цепи мРНК, участок siRNA может с помощью комплекса ферментов, называемого DICER, сначала достроить вторую цепочку мРНК, а затем разрезать ее в разных местах, создавая таким образом разнообразные "вторичные" siRNA. Они в свою очередь формируют RISC и проводят мРНК через все стадии, о которых шла речь выше, вплоть до ее полного уничтожения. Такие "вторичные" молекулы смогут специфично связываться не только с тем участком вирусной мРНК, к которому была направлена "первичная" молекула, но также и с другими участками, что резко усиливает эффективность клеточной защиты.
У млекопитающих в отличие от насекомых и растений работает и другая система защиты. При попадании в "зрелую" (дифференцированную) клетку млекопитающего чужой РНК, длина которой больше 30 нуклеотидов, клетка начинает синтез интерферона (ряд белков со сходными свойствами, выделяемые клетками организма в ответ на вторжение вируса). Интерферон, связываясь со специфическими рецепторами на клеточной поверхности, способен стимулировать целую группу генов. В результате в клетке синтезируется несколько видов ферментов, которые тормозят синтез белков и расщепляют вирусные РНК. Кроме того, интерферон может действовать и на соседние, еще не зараженные клетки, блокируя тем самым возможное распространение вируса.
Как можно заметить, обе системы во многом схожи. Даже названия "interferon" и "(RNA) interference" происходят от общего корня. Но есть у них и одно очень существенное различие: если интерферон при первых признаках вторжения просто "замораживает" работу клетки, не позволяя (на всякий случай) производство многих белков в клетке, то система siRNA отличается чрезвычайной разборчивостью: каждая siRNA будет распознавать и уничтожать только свою, специфическую мРНК. Замена всего лишь одного нуклеотида внутри siRNA ведет к резкому снижению эффекта интерференции.
Функциональная роль РНК-интерференции.
Итак, мы видим, что основная "специальность" siRNA в клетке – это блокирование генов, участок которых соответствует одной из цепочек внутри siRNA. Но почему механизм РНК-интерференции существует в клетках? Какие процессы он призван регулировать? Что из себя представляют те мРНК, на которые направлено "жало" малых РНК, и почему их необходимо уничтожать?
Одно из возможных объяснений роли РНК-интерференции – это защита организмов от РНК-содержащих вирусов и мобильных элементов, перемещающихся посредством РНК. Таким образом, двухцепочеченые РНК (дцРНК) могут быть важными звеньями иммунной системы, распознающими и ликвидирующими чужую РНК. В случае, если в клетку проник РНК-содержащий вирус, такая система защиты не даст ему размножиться. Если же вирус содержит ДНК, дцРНК будет мешать ему производить вирусные белки.
Как предполагают, роль коротких РНК в клетке не ограничивается только защитой от вирусов и других чужеродных РНК. Дело в том, что геном любого многоклеточного организма включает множество элементов, которые когда-то были привнесены в него в процессе эволюции извне, например как результат встраивания вируса. Из всего материала, содержащегося в хромосомах человека, 34% приходится на долю элементов, называемых LINEs и SINEs (соответственно, Long и Short Interspersed Nuclear Elements), о функциях которых известно только то, что они могут время от времени копировать себя и перемещаться с одного места хромосомы на другое. Те участки ДНК, которые достались нам от ретровирусов (8% генома) и транспозоны (3%) также способны менять свое место в геноме. LINEs, SINEs, остатки вирусной ДНК и транспозоны, за их способность к перемещениям названные подвижными, или мобильными, элементами генома, представляют значительную опасность для наших хромосом. Некоторые из них – остатки вирусов, или протоонкогены – способны при "включении" вызывать рак, другие мобильные элементы, размножаясь и перемещаясь, меняют структуру хромосом, что может привести к мутациям.
Например, у плодовой мушки дрозофилы более 80% спонтанных мутаций возникают именно из-за "хулиганского" поведения ее собственных мобильных элементов. Их перемещения внутри генома так индивидуальны и непредсказуемы, что положение некоторых из них может служить "молекулярным паспортом", точно определяющим личность хозяина, что уже используется на практике. Понятно, что каждому организму было необходимо разработать систему жесткого контроля за поведением мобильных элементов. Как раз эту "надзирательскую" функцию, как полагают, и выполняет система внутриклеточных siRNA. На модели все того же C. elegans, например, было показано, что отключение генов, кодирующих некоторые из малых РНК, ведет к активизации перемещений мобильных элементов в его хромосомах и соответственно – к повышению уровня мутаций.
Кроме того, ошибки в развитии органов и тканей при отключении генов, кодирующих систему siRNA у подопытных животных, а также ее активность в "незрелых" клетках указывают на то, что механизм РНК-интерференции активно участвует в регуляции программы "созревания" клеток и как следствие может играть одну из ключевых ролей в формировании целостного организма.
Еще одна из предполагаемых нормальных функций siRNA – отслеживание неправильно обработанных копий других типов РНК в клетке.
Наконец, в 2002 г. исследователи неожиданно выяснили, что действие siRNA может не ограничиваться лишь временным выключением генов на уровне РНК. Имеются показания, что в некоторых случаях siRNA, видимо, воздействует прямо на ДНК, изменяя структуру хроматина и способствуя длительному "замолканию" одних и, возможно, активизации других генов.
Круговорот энергии в экосистемах, возобновляемые и не возобновляемые источники энергии.
П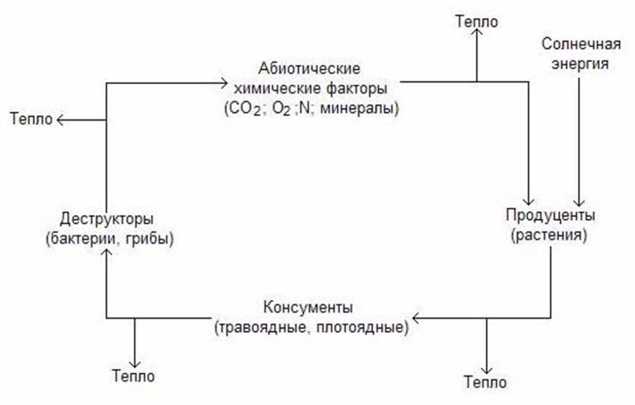 ищевые
цепи и пищевая сеть
ищевые
цепи и пищевая сеть
В функционирующей природной экосистеме не существует отходов. Все организмы, живые или мертвые, потенциально являются пищей для других организмов: гусеница ест листву, дрозд питается гусеницами, ястреб способен съесть дрозда. Когда растения, гусеница, дрозд и ястреб погибают, они в свою очередь перерабатываются редуцентами.
Пищевая цепь – последовательность организмов, в которой каждый из них съедает или разлагает другой. Пищевые цепи – это также движение питательных веществ от продуцентов, консументов (травоядных, плотоядных и всеядных) к редуцентам и обратно к продуцентам. Все организмы, пользующиеся одним типом пищи, принадлежат к одному трофическому уровню (от греческого слова trophos – «питающиеся»).
Организмы природных экосистем вовлечены в сложную сеть многих связанных между собой пищевых цепей. Такая сеть называется пищевой сетью.
Д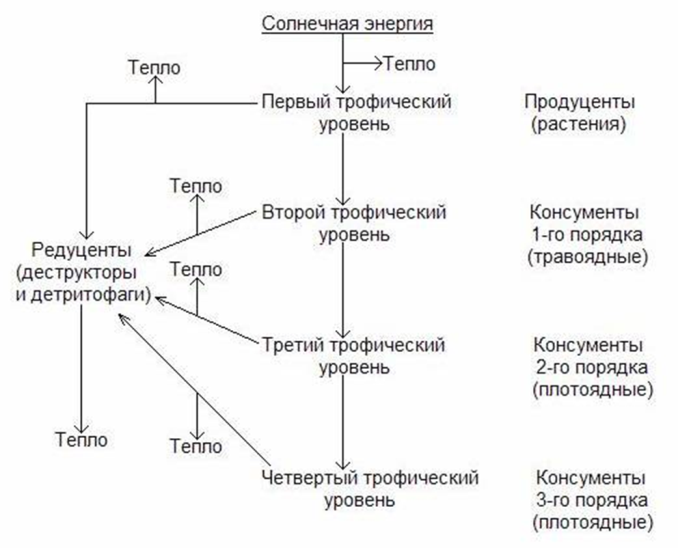 вижение
энергии в экосистемах происходит
посредством двух связанных типов пищевых
сетей: пастбищной и детритной.
вижение
энергии в экосистемах происходит
посредством двух связанных типов пищевых
сетей: пастбищной и детритной.
В пастбищной пищевой сети живые растения поедаются фитофагами, а сами фитофаги являются пищей для хищников и паразитов.
В детритной пищевой сети отходы жизнедеятельности и мертвые организмы разлагаются детритофагами и деструкторами до простых неорганических соединений, которые вновь используются растениями.
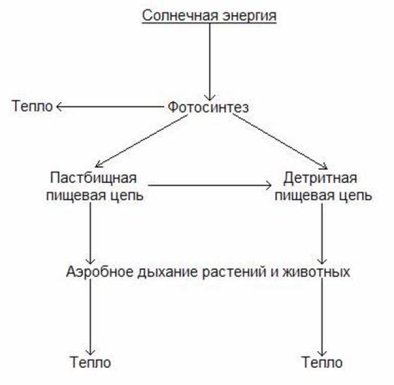
Пирамиды энергетических потоков
С каждым переходом из одного трофического уровня в другой в пределах пищевой цепи или сети совершается работа и в окружающую среду выделяется тепловая энергия, а количество энергии высокого качества, используемой организмами следующего трофического уровня, снижается.
Правило 10% (правило Линдемана)
При переходе с одного трофического уровня на другой 90% энергии теряется, и 10% передается на следующий уровень. Чем длиннее пищевая цепь, тем больше теряется полезной энергии. Поэтому длина пищевой цепи обычно не превышает 4-5 звеньев.
Пирамиды численностей и биомасс
Можно собрать все образцы организмов в экосистеме и подсчитать численность всех видов, обнаруженных на каждом трофическом уровне. Такая информация необходима для создания пирамиды численностей для экосистем.

Обобщенные пирамиды численностей в экосистемах
Сухой вес всех органических веществ, содержащихся в организмах экосистемы, называется биомассой. Каждый трофический уровень пищевой цепи или сети содержит определенное количество биомассы. Ее можно вычислить, если собрать все живые организмы с различных произвольно выбранных участков. Собранные экземпляры необходимо рассортировать по трофическим уровням, высушить и взвесить. Полученные данные в дальнейшем используются для построения пирамиды биомасс для определенной экосистемы.
 Обобщенные
пирамиды биомасс в экосистемах. Размер
каждого слоя пропорционален сухой массе
на квадратный метр всех организмов на
данном трофическом уровне
Обобщенные
пирамиды биомасс в экосистемах. Размер
каждого слоя пропорционален сухой массе
на квадратный метр всех организмов на
данном трофическом уровне
Биологической продукцией называют то количество органического вещества, которое создается в единицу времени на определенном трофическом уровне. Различают первичную и вторичную биологическую продукцию. Первичную создают растения из неорганических компонентов. Валовая первичная продукция определяется скоростью фотосинтеза, которая зависит и от вида растений, и от условий среды. Значительную часть валовой первичной продукции (в среднем 50–60%) растения тратят на поддержание собственной жизнедеятельности, а оставшаяся часть идет на рост – увеличение массы. Эту часть, заключенную в созданной биомассе, называют чистой первичной продукцией. За счет чистой первичной продукции в конечном счете и существуют все гетеротрофные организмы биоценоза.
Вторичная биологическая продукция – это органическое вещество, создаваемое гетеротрофами, т.е. переработчиками. Она возникает за счет первичной чистой продукции и, по правилу передачи энергии по цепям питания, составляет не более 10% от растительной. Таким образом, в природе на создание 1 кг массы травоядного животного тратится в 10 раз больше солнечной энергии, чем на создание 1 кг массы растений, а на 1 кг массы хищника – по крайней мере в 100 раз.
В сельском хозяйстве законы передачи энергии по цепям питания выражаются в том известном всем обстоятельстве, что 1 кг пшеницы вырастить гораздо дешевле, чем получить 1 кг говядины или свинины. По этой же причине на мясо не разводят хищных животных, а среди одомашненных растительноядных для этой цели используют в основном тех, которые наиболее эффективно перерабатывают пищу на собственный прирост: кур, индеек, уток, свиней, крупный и мелкий рогатый скот и т.п. Как уже сказано, 10% – это усредненная величина, а конкретные живые организмы могут характеризоваться разной величиной этого показателя. Среди сельскохозяйственных животных, у свиней например, КПД использования 1 кг кормов на прирост 1 кг живой массы может достигать 20% и более. Для других видов, несмотря на старания селекционеров, достичь таких результатов не удается. Фазанов и павлинов разводят почти исключительно в декоративных целях, поскольку доля энергии, использующаяся на прирост, у них составляет всего 2–3%.
Эффективность функционирования начальных звеньев пищевой цепи
В каждую экосистему входят группы организмов разных видов, различимые по способу питания:
автотрофы;
гетеротрофы;
консументы – потребители органического вещества живых организмов;
детритофаги, или сапрофаги, - организмы, питающиеся мертвым органическим веществом – остатками растений и животных;
редуценты – бактерии и низшие грибы – завершают деструктивную работу консументов и сапрофагов, доводя разложение органики до ее полной минерализации и возвращая в среду экосистемы последние порции двуокиси углерода, воды и минеральных элементов.
Все эти группы организмов в любой экосистеме тесно взаимодействуют между собой, согласуя потоки вещества и энергии.
Прослеживая пищевые взаимоотношения между членами биоценоза, можно построить пищевые цепи и пищевые сети питания различных организмов. Различают несколько типов пищевых систем:
пастбищные пищевые цепи (цепи эксплуататоров);
цепи паразитов;
детритные цепи.
Благодаря определенной последовательности пищевых отношений различаются отдельные трофические уровни переноса веществ и энергии в экосистеме, связанные с питанием определенной группы организмов. Так, первый трофический уровень во всех экосистемах образуют продуценты – растения, второй – первичные консументы- фитофаги, третий – вторичные консументы- зоофаги и т.д.
Вход системы – поток солнечной энергии. Часть энергии, поглощенная на первом трофическом уровне растениями, преобразуется фотосинтезом в энергию химических связей углеводов и других органических веществ. Это валовая первичная продукция (ВПП, брутто-продукция) экосистемы. Часть ее веществ окисляется в процессе дыхания растений и освобождает энергию. Эта энергия используется в других биохимических процессах в растении и в конечном счете также рассеивается в виде тепла. Оставшаяся часть новообразованных органических веществ обусловливает прирост биомассы растений – чистую первичную продукцию (ЧПП, нетто-продукцию) экосистемы. Отношение чистой продукции к валовой (коэффициент эффективности фотосинтеза) зависит от типа растительности.
“Правило 1%”: для биосферы в целом доля возможного потребления чистой первичной продукции (на уровне консументов высших порядков) не превышает 1%.
Все природные ресурсы по исчерпаемости делятся на две группы: исчерпаемые и неисчерпаемые.
I. Исчерпаемые ресурсы. Они образуются в земной коре или ландшафтной сфере, но объемы и скорости их формирования измеряются по геологической шкале времени. В то же время потребности в таких ресурсах со стороны производства или для организации благоприятных условий обитания человеческого общества значительно превышают объемы и скорости естественного восполнения. В результате неизбежно наступает истощение запасов природного ресурса. В группу исчерпаемых включены ресурсы с неодинаковыми скоростями и объемами формирования. Это позволяет провести их дополнительную дифференциацию.
На основе интенсивности и скорости естественного образования исчерпаемые ресурсы делят на подгруппы:
1. Невозобновляемые, к которым относят:
а) все виды минеральных ресурсов или полезные ископаемые. Они постоянно образуются в недрах земной коры в результате непрерывно протекающего процесса рудообразования, но масштабы их накопления столь незначительны, а скорости образования измеряются многими десятками и сотнями миллионов лет (например, возраст каменных углей насчитывает более 350 млн. лет), что практически их учитывать в хозяйственных расчетах нельзя. Освоение минерального сырья происходит по исторической шкале времени и характеризуется всевозрастающими объемами изъятия. В этой связи все минеральные ресурсы рассматриваются в качестве не только исчерпаемых, но и невозобновляемых.
б) земельные ресурсы в их естественном природном виде - это материальный базис, на котором происходит жизнедеятельность человеческого общества. Морфологическое устройство поверхности (т. е. рельеф) существенно влияет на хозяйственную деятельность, на возможность освоения территории. Однажды нарушенные земли (например, карьерами) при крупном промышленном или гражданском строительстве в своем естественном виде уже не восстанавливаются.
2. Возобновляемые ресурсы, к которым принадлежат:
а) ресурсы растительного и
б) животного мира.
И те и другие восстанавливаются довольно быстро, и объемы естественного возобновления хорошо и точно рассчитываются. Поэтому при организации хозяйственного использования накопленных запасов древесины в лесах, травостоя на лугах или пастбищах, промысла диких животных в пределах, не превышающих ежегодное возобновление, можно полностью избежать истощения ресурсов.
3. Относительно (не полностью) возобновляемые. Некоторые ресурсы хотя и восстанавливаются в исторические отрезки времени, но возобновляемые объемы их значительно меньше объемов хозяйственного потребления. Именно поэтому такие виды ресурсов оказываются весьма уязвимыми и требуют особенно тщательного контроля со стороны человека. К относительно возобновляемым ресурсам относятся и очень дефицитные природные богатства:
а) продуктивные пахотно-пригодные почвы;
б) леса с древостоями спелого возраста;
в) водные ресурсы в региональном аспекте.
Продуктивных пахотно-пригодных почв сравнительно немного (по разным оценкам их площадь не превышает 1,5-2,5 млрд. га). Наиболее продуктивные почвы, относящиеся к первому классу плодородия, занимают, по оценкам ФАО, всего 400 млн. га. Продуктивные почвы образуются крайне медленно – на формирование 1 мм слоя, например, черноземных почв требуется более 100 лет.
II. Неисчерпаемые ресурсы. Среди тел и явлений природы ресурсного значения имеются и такие, которые практически неисчерпаемы, К ним относятся климатические и водные ресурсы.
А) Климатические ресурсы. Наиболее жесткие требования к климату предъявляют сельское хозяйство, рекреационное и лесное хозяйство, промышленное и гражданское строительство и др. Обычно под климатическими ресурсами понимают запасы тепла и влаги, которыми располагает конкретная местность или регион. Общие запасы тепла, поступающие за год на 1 м.кв. поверхности планеты, равны 3.16 х 10 Дж (радиационный бюджет в среднем для планеты). Территориально и по сезонам года тепло распределяется неравномерно, хотя в среднем для Земли температура воздуха равна примерно + 15°С. Суша в целом неплохо обеспечена и атмосферной влагой: на ее поверхность ежегодно выпадает в среднем около 119 тыс. куб. км осадков. Но распределяются они еще более неравномерно, чем тепло, и в пространственном и во временном отношениях На суше известны районы, получающие ежегодно более 12000 мм осадков, к обширные местности, где за год выпадает менее 50-100 мм. В среднем многолетнем выражении и запасы тепла и объемы выпадающей атмосферной влаги довольно постоянны, хотя от года к году могут наблюдаться существенные колебания в обеспечении территории теплом и влагой. Так как эти ресурсы формируются в определенных звеньях теплового и водного круговоротов, постоянно действующих над планетой в целом и над ее отдельными регионами, запасы тепла и влаги могут рассматриваться как неиссякаемые в определенных количественных пределах, точно установленных для каждого района.
Б) Водные ресурсы. Земля обладает колоссальным объемом воды - около 1,5 млрд. куб. км. Однако 98% этого объема составляют соленые воды Мирового океана, и только 28 млн. куб. км - пресные воды. Поскольку уже известны технологии опреснения соленых морских вод, воды Мирового океана и соленых озер можно рассматривать как потенциальные водные ресурсы, использование которых в будущем вполне возможно. Мировое хозяйство расходует для своих нужд около 4-4,5 тыс. куб. км, что равно примерно 10% общего водозапаса, и, следовательно, при условии соблюдения принципов рационального водопользования эти ресурсы можно рассматривать как неисчерпаемые. Однако при нарушении этих принципов ситуация может резко обостриться, и даже в планетарном масштабе может ощущаться дефицит чистых пресных вод. А пока природная среда ежегодно "дарит" человечеству в 10 раз больше воды, чем ему нужно для удовлетворения самых разнообразных потребностей.
Ресурсы - все извлекаемое нами из природной среды для удовлетворения своих потребностей и желаний (чистый воздух, питьевая вода, нефть, железная руда, древесина, культурные растения). Большая часть ресурсов требует для своего употребления определенных условий, определенного развития технологии (например, нефть, никелевые руды, бокситы). Значит, все, что угодно может стать ресурсом при соответствующих технологии и потребностях.
Типы ресурсов:
исчерпаемые (возобновимые и невозобновимые);
неисчерпаемые (солнечная энергия, энергия ветра, приливов, геотермальная энергия и т.д.
Невозобновимые ресурсы существуют в ограниченных количествах (запасах) в различных частях земной коры. Примерами являются медь, алюминий, уголь и нефть. Невозобновимые ресурсы считаются экономически истощенными, когда выработаны 80% их оцененных запасов. По достижении этого предела разведка, добыча и переработка остающихся запасов обходится дороже розничной цены. Некоторые виды невозобновимых ресурсов могут использоваться повторно или циклически, увеличивая, таким образом, потенциал использования ресурсов алюминия, меди, железа или, например, стекла.
Потенциально возобновимые - ресурсы, запасы которых хотя и могут быть истощены или загрязнены в результате слишком быстрого потребления, однако в нормальных условиях восстановятся в результате естественных процессов (деревья, пресные воды рек и озер, почвы, дикие животные).
Билет № 12
Экологические проблемы в Беларуси.
Экологическая ситуация на территории Беларуси в последние годы оставалась относительно благополучной. Высокие темпы экономического роста не сопровождались усилением воздействия на окружающую среду.
В структуре земельного фонда продолжились благоприятные в экологическом отношении изменения, связанные со снижением доли пахотных и увеличением доли лесных земель. Общая площадь, занятая природными экосистемами (лесными, луговыми, болотными и водными), в два раза превысила таковую, занятую угодьями, преобразованными хозяйственной деятельностью (пашней, застроенными и нарушенными землями). Данное соотношение естественных и измененных хозяйственной деятельностью природных комплексов способствует сохранению экологического равновесия, а также ландшафтного и биологического разнообразия.
Вместе с тем в рассматриваемом году продолжали действовать факторы, вызывающие проблемные ситуации, связанные с загрязнением окружающей среды и деградацией природно-ресурсного потенциала. Они обусловлены, во-первых, функционированием национальной экономики и, в первую очередь, производственного комплекса, во-вторых, трансграничным переносом загрязняющих веществ, в-третьих, наличием на территории страны унаследованных проблем, не решенных в прошлом.
Приоритетные экологические проблемы страны включают в себя радиоактивное загрязнение территории, загрязнение атмосферного воздуха, загрязнение поверхностных и подземных вод, загрязнение и деградацию почв, образование и накопление отходов. При их характеристике использовались данные Национальной системы мониторинга окружающей среды Республики Беларусь.