

Частина 3. Біохімія фізіологічних функцій
РОЗДІЛ 13. БІОХІМІЯ М’ЯЗОВОЇ СИСТЕМИ
13.1. Теоретична частина
Рухова активність є основою усіх пристосувальних реакцій людини і тварин до змін зовнішнього середовища. Будь-який рух людини відбувається в результаті скорочення органів руху і сили – м’язів. М’язова тканина займає перше місце за об’ємом серед інших тканин людини. В організмі дорослого чоловіка на м’язи припадає 40-45% маси тіла (у жінок того ж віку маса м’язів зазвичай менша, з чим і пов’язані статеві відмінності в проявах м’язової сили і рівня фізичної працездатності), у людей похилого віку – 30%, дітей – близько 25%. У спортсменів силовиків маса м’язів може досягати 50-55%, а у культуристів – 60-70% загальної маси тіла.
М’язи – основний біохімічний перетворювач потенціальної (хімічної) енергії в кінетичну (механічну). Ефективне перетворення хімічної енергії в механічну можливе при дотриманні певних умов:
− сталого забезпечення хімічною енергією (в м’язах хребетних хімічна енергія зосереджена в молекулах АТФ і креатинфосфату);
− існування засобів регуляції механічної активності (швидкості, тривалості і сили скорочення);
− забезпечення нервовою системою контролю процесу перетворення;
− забезпечення механізму повернення системи у вихідний стан.
За своїми властивостями м’язи характеризуються великою еластичністю, пластичністю та скоротливістю.
13.1.1. М’язові волокна та їх структурна організація
В організмі хребетних існує три типи м’язів: скелетні, серцеві (міокард) та гладкі. Вони відрізняються морфологічно, біохімічно і функціонально. Скелетні та серцеві м’язи мають поперечну смугастість; в гладких м’язах вона відсутня. Скелетні м’язи функціонують довільно, серцеві і гладкі – мимовільно. Скелетні м’язи прикріплені в основному до кісток, що і обумовлює їх назву.
Для розуміння біохімії м’язової діяльності найбільший інтерес представляє функціонування скелетних м’язів.
Окремі м’язи різних частин тіла оточені оболонкою сполучної тканини і мають складну морфологічну будову. Кожний м’яз складається з пучка м’язових волокон, які містять багаточисельні скоротливі нитки – міофібрили (рис. 13.1).
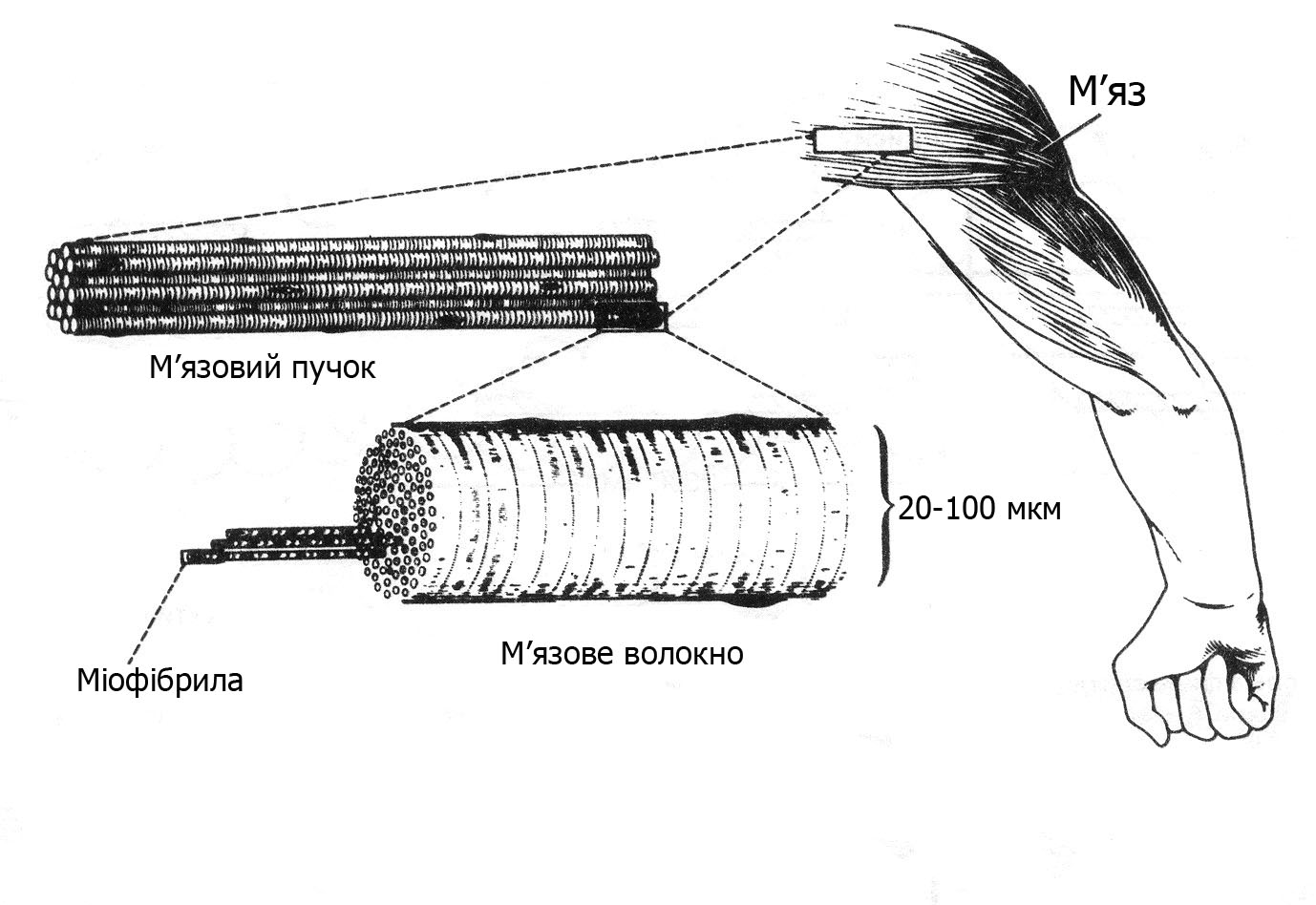
Рис. 13.1. Структурна організація скелетних м’язів людини
М’язове волокно − структурна одиниця скелетних м’язів, багатоядерна циліндричної форми видовжена поперечносмугаста клітина. Кожне м’язове волокно має велику кількість (1 тис. і більше) м’язових ниток – міофібрил. У різних людей в одних і тих же м’язах може бути різна кількість волокон, що впливає на їх силові властивості, процеси адаптації до м’язової роботи. Чим більше в м’язах волокон, тим вища можливість прояву максимальної сили м’язів. Товщина окремих м’язових волокон – 10-100 мкм, довжина – від 1-2 до 10-12 см. При народженні дитини товщина м’язових волокон становить близько 1/5 товщини волокон дорослих людей. Діаметр м’язових волокон значно зростає під впливом систематичних фізичних навантажень.
В скелетних м’язах є кілька типів м’язових волокон, які відрізняються скоротливими і метаболічними властивостями. До основних типів волокон відносяться червоні, білі та проміжні волокна. Саркоплазма червоних волокон містить багато міоглобіну, який і надає їм відповідного забарвлення, а також численні мітохондрії. М’язи, в яких переважають червоні волокна, скорочуються повільніше, але довго і без ознак втоми. Білі волокна містять менше міоглобіну та мітохондрій, але більше глікогену і глюкози. В білих волокнах вища АТФ-азна активність міозину. М’язи, що складаються здебільшого з білих волокон, швидко переходять від стану спокою до максимальної активності, скорочуються значно швидше, але раніше втомлюються, оскільки вичерпуються запаси глікогену, а глюкоза з крові надходить повільно.
М’язове волокно покрите тонкою еластичною мембраною − сарколемою (від грец. sarkós − м’ясо, плоть і lemma − шкірка, шкаралупа). Сарколема відіграє важливу роль у виникненні і проведенні збудження. Вона володіє вибірковою проникністю для різних речовин і має транспортні системи за допомогою яких підтримується різна концентрація йонів Na+, К+, Cl- всередині клітини і в міжклітинній рідині, що призводить до виникнення на її поверхні мембранного потенціалу. Утворення мембранного потенціалу дії під впливом нервового імпульсу – необхідна умова виникнення збудження м’язового волокна.
Внутрішній простір м’язового волокна заповнений внутрішньоклітинною рідиною − саркоплазмою. До її складу входить саркоплазматичний матрикс і ретикулум.
Саркоплазматичний матрикс – це рідина, в яку занурені скоротливі елементи м’язового волокна – міофібрили. Крім міофібрил до складу матрикса входять гранули глікогену, крапельки жиру, фосфатні речовини та інші малі молекули і йони, pозчинні білки (16,5–21,0 %).
В кожному м’язовому волокні знаходиться до 1000 і більше міофібрил. Це спеціалізовані скоротливі структури діаметром 1-3 мкм. До складу міофібрил входять товсті міозинові і тонкі актинові філаменти.
Просторове розташування міофіламентів визначає посмугованість скелетних м’язів, яку добре видно під мікроскопом. Тонкі Z-мембрани (телофрагма-Т), до яких прикріпляються тонкі філаменти, ділять міофібрили на саркомери. Саркомер – це основна структурно-функціональна скоротлива одиниця міофібрили (рис. 13.2). В саркомерах актинові філаменти розташовані між міозиновими, які мають довгі «хвости» і «головки». Головки міозинових молекул повернуті в косому напрямі до актинових ниток і мають назву поперечних містків.
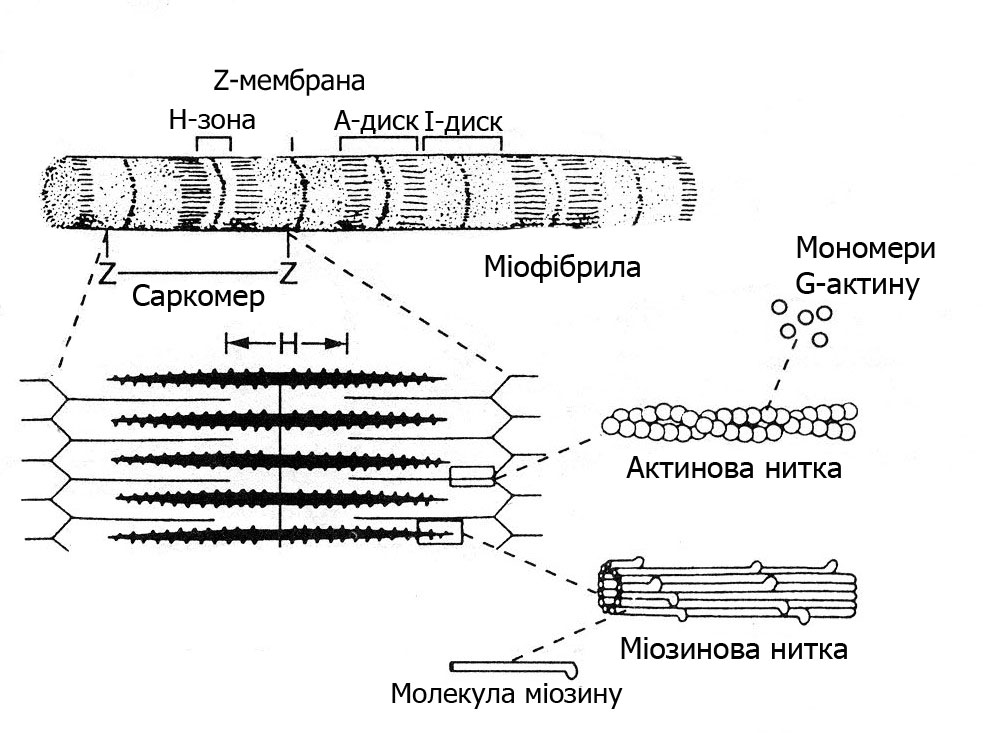
Рис. 13.2. Будова саркомера
Поперечна посмугованість міофібрил є наслідком чергування ділянок саркомера з сильним і слабким заломленням променів. В тій частині саркомера, де розміщені тільки тонкі нитки (диск І), заломлення променів невелике. В місцях перекриття товстих і тонких ниток (диск А) заломлення променів подвійне, а тому вони виглядають темними. Всередині диска А тонких ниток немає – це Н-зона, вона світла. Посередині диска А поперек проходить мембрана – ізофрагма (М), вона взаємозв’язана з телофрагмою (Z-лінією) і з сарколемою.
Міофібрили згруповані в пучки (колонки) по 4-10 штук в кожному. Колонки тісно зв’язані з транспортною системою м’язового волокна (саркоплазматичною сіткою), по якій циркулюють поживні речовини і розповсюджується електрична хвиля збудження.
Саркоплазматичний ретикулум (СР) − це внутрішньоклітинна мембранна система, яка складається з поперечних (Т-трубочок) і поздовжніх трубочок, розташованих між міофібрилами паралельно до них (рис. 13.3). На внутрішній мембрані розташовані білки, які здатні зв’язувати йони Са2+. Основна функція СР – регуляція йонів Кальцію в просторі між актином та міозином, рівень якого змінюється при скороченні-розслаблені від 10-5 до 10-7 моль/л. Під дією нервового імпульсу СР виділяє йони Са2+, а після припинення його дії знову поглинає. На мембрані СР розміщені також рибосоми, де проходить біосинтез білків.
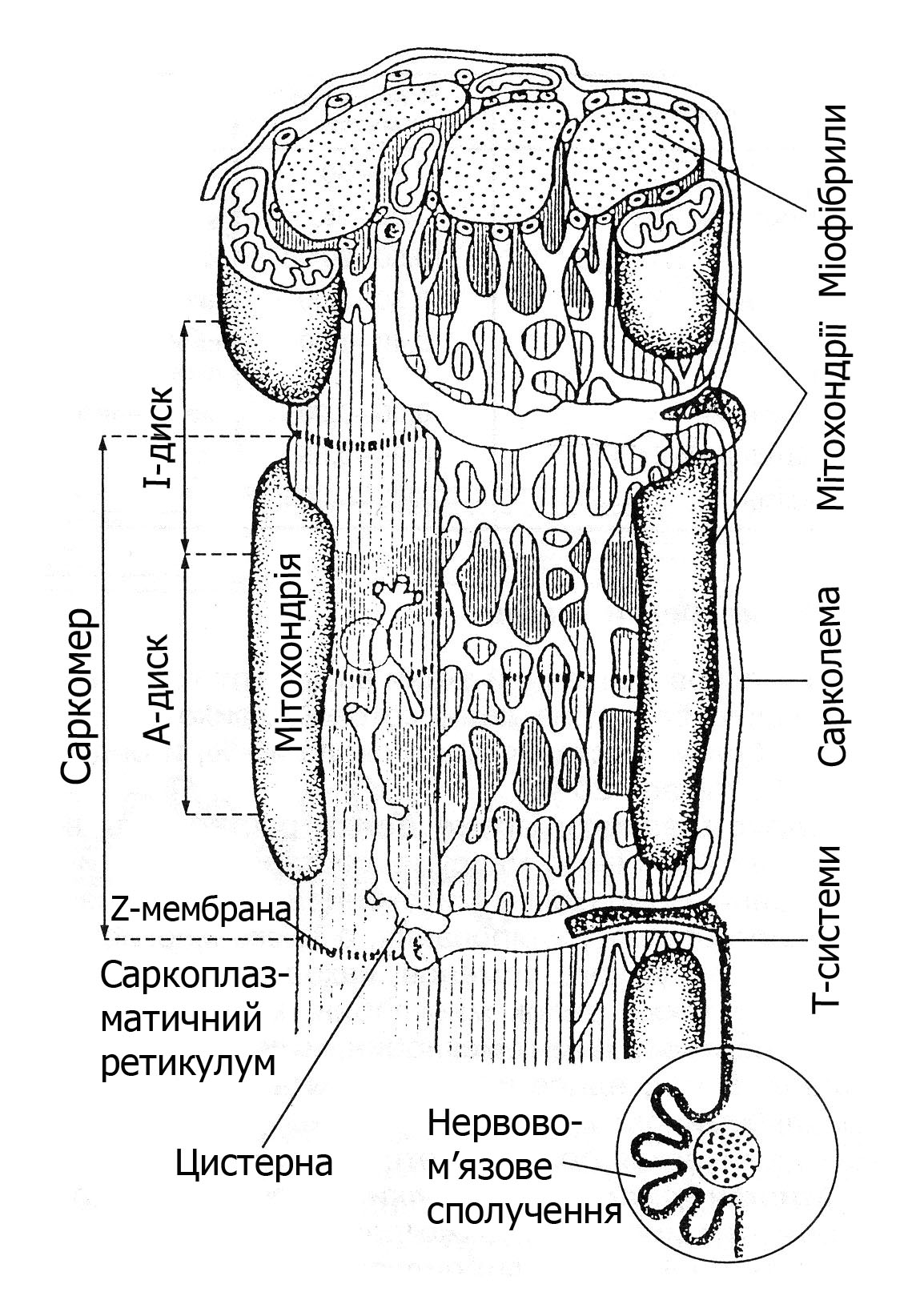
Рис. 13.3. Схема розміщення Т-систем
і саркоплазматичного ретикулуму у м’язовому волокні
В м’язовій клітині по ходу міофібрил знаходяться мітохондрії (саркосоми). Мітохондрії виконують функції «енергетичних станцій» м’язового волокна, оскільки в них утворюється АТФ – енергія для м’язового скорочення. Їх кількість зумовлює функціональну активність м’язів. До складу мітохондрій входять білки, жири, вуглеводи, нуклеїнові кислоти, а також велика кількість ферментів, які беруть безпосередню участь у процесах ресинтезу АТФ. Найбільше мітохондрій в постійно працюючому серцевому м’язі.
Непосмугована м’язова тканина. Гладенькі м’язи утворюють стінки внутрішніх органів (органів травної системи, бронхів, кровоносних і лімфатичних судин, сечового міхура, матки, тощо), шкіри і залоз. На відміну від посмугованої м’язової тканини міофібрили гладеньких м’язів не мають посмугованості. До їх складу входить білок тономіозин (різновидність актоміозину), а також леатонін, функцією якого є підтримання тривалого тонічного напруження. Цьому сприяє хаотичне розташування скоротливих білків у волокнах гладеньких м’язів. Довжина клітин гладеньких м’язів залежить від органу, до складу якого вони входять (20-500 мкм), діаметр – 5-20 мкм.
Гладенькі м’язи менш збудливі, ніж поперечносмугасті, вони мають більш тривалий рефрактерний період і невелику швидкість поширення нервового імпульсу (2-15 см/с). Збудження по непосмугованих м’язах може вільно передаватися з одного волокна на інше. Ці та інші морфофізіологічні особливості гладеньких м’язів зумовлюють ряд відмінних ознак їх функціонування.