

10.3.1. Общая характеристика
Нуклеиновые кислоты (полинуклеотиды) — это биополимеры, структурными единицами которых являются нуклеотиды, состоящие из азотистого основания, моносахарида (рибозы или дезоксирибозы) и фосфорной кислоты.
Основная биологическая функция нуклеиновых кислот — хранение и передача наследственной информации. Нуклеиновые кислоты служат материальными носителями генетического кода, который определяет аминокислотную последовательность в белках. Именно эта генетическая информация программирует структуру и метаболическую активность живых организмов. Нуклеиновые кислоты содержатся во всех живых организмах и играют исключительно важную роль в биосинтезе белков. Структурные компоненты также выполняют функции кофакторов, аллостерических эффекторов; входят в состав коферментов.
В организме нуклеиновые кислоты находятся в основном в составе смешанных биополимеров — нуклеопротеинов. Как следует из названия, структурными компонентами таких образований являются нуклеиновые кислоты и белки.
Нуклеиновые основания являются производными азотсодержащих гетероциклических соединений — пиримидина или пурина. В состав полинуклеотидных цепей ДНК входят пиримидиновые основания тимин (Т) и цитозин (Ц); в случае РНК — урацил (У) и цитозин. Данные соединения обнаруживают способность к лак- тим-лактамной таутомерии:
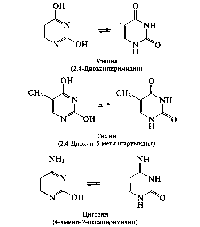
Аденин (А) и гуанин (Г) являются производными пурина и называются пуриновыми нуклеиновыми основаниями:

Следует отметить, что пиримидиновые основания входят в состав полинуклеотидов в лактамной форме, что обусловлено возможностью образования водородных связей между остатками оснований в цепях нуклеиновых кислот: аденин—тимин и гуанин— цитозин в ДНК; аденин—урацил и гуанин—цитозин в РНК (рис. 10.4).
Рассмотрим закономерности состава и количественного содержания азотистых оснований в нуклеиновых кислотах (ДНК):
мольная доля пуриновых оснований (аденина и гуанина) равна мольной доле пиримидиновых оснований (цитозина и тимина);
суммарное количество вещества аденина и цитозина равно суммарному количеству вещества гуанина и тимина;
количества вещества комплементарных друг другу оснований (аденина и тимина, гуанина и цитозина) равны;
коэффициент специфичности для животных, равный отношению суммы количества вещества гуанина и цитозина к сумме количества вещества аденина и тимина, меньше единицы.
Рис.
10.4. Водородные связи в паре оснований
аденин—тимин (а) и гуанин-цитозин
(б)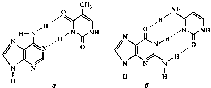
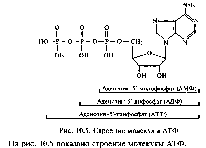
Уровни структурной организации нуклеиновых кислот
Последовательность мононуклеотидов в полинуклеотидной цепи называют первичной структурой (рис. 10.6). Нуклеотиды связаны между собой посредством 3',5'-фосфодиэфирных связей. Как правило, молекулы рибонуклеиновых кислот (РНК) состоят из одной полинуклеотидной цепи, тогда как большинство молекул ДНК двухцепочечные.

Две цепи ДНК, связанные водородными связями между парами комплементарных оснований, образуют правовращающую спираль. Таким образом возникает вторичная структура. На следующем этапе компактизации полинуклеотидной цепи на некоторых участках двойной спирали ДНК могут образоваться «суперспирали» или кольцевые формы за счет сил Ван-дер-Ваальса — возникает третичная структура ДНК.
В составе молекулы ДНК выделено значительно большее число нуклеотидных остатков, чем в молекуле РНК; ДНК имеет молекулярную массу порядка 107 и в условиях клетки нерастворима.
Существует несколько разновидностей РНК, среди которых наиболее активными в биосинтезе белка являются:
транспортная РНК молекулярной массой -25000;
информационная, или матричная, РНК молекулярной массой 105—106.
Длина молекул ДНК человека составляет порядка 3 — 5 мм, молекула РНК значительно короче — до 0,1 мм.
М акромолекулы
ДНК связаны между собой попарно при
помощи водородных связей в виде двойной
спирали постоянного диаметра (рис.
10.7). Остатки нуклеиновых оснований
направлены внутрь спирали, диаметр
которой равен примерно 2 нм. На один
виток спирали приходится 10 пар оснований.
Для обеспечения наибольшей устойчивости
этой структуры необходимо, чтобы
число водородных связей было максимально
возможным. Это достигается определенным
соответствием в расположении остатков
оснований одной спирали по отношению
к остаткам другой: тиминовые группы
располагаются напротив адениновых,
цитозиновые — напротив гуаниновых и
т.д. Кроме того, только при выполнении
этого условия обеспечивается
экспериментально доказанное постоянство
суммарных размеров боковых групп и
неизменность диаметра двойной
спирали на всем ее протяжении. В этой
взаимной обусловленности порядка
чередования звеньев в обеих цепях
заключается принцип
комп- лементарности.
акромолекулы
ДНК связаны между собой попарно при
помощи водородных связей в виде двойной
спирали постоянного диаметра (рис.
10.7). Остатки нуклеиновых оснований
направлены внутрь спирали, диаметр
которой равен примерно 2 нм. На один
виток спирали приходится 10 пар оснований.
Для обеспечения наибольшей устойчивости
этой структуры необходимо, чтобы
число водородных связей было максимально
возможным. Это достигается определенным
соответствием в расположении остатков
оснований одной спирали по отношению
к остаткам другой: тиминовые группы
располагаются напротив адениновых,
цитозиновые — напротив гуаниновых и
т.д. Кроме того, только при выполнении
этого условия обеспечивается
экспериментально доказанное постоянство
суммарных размеров боковых групп и
неизменность диаметра двойной
спирали на всем ее протяжении. В этой
взаимной обусловленности порядка
чередования звеньев в обеих цепях
заключается принцип
комп- лементарности.
Рис. 10.7. Двойная спираль ДНК
Спирализация приводит к возникновению вторичной структуры ДНК. Экспериментально было установлено, что чем больше в молекуле ДНК остатков аденозина и тимина, тем при более низкой температуре происходит разрушение вторичной структуры, т.е. раскручивание спирали. Это объясняется тем, что суммарный эффект связывания аденин—тимин (две водородные связи) менее выражен, чем аналогичный эффект для пары гуанин—цитозин (три водородные связи).
Как было сказано, вторичная структура и ДНК, и РНК в значительной степени определяется водородными связями между парами комплементарных азотистых оснований, поэтому любые воздействия, способные разрушать водородные связи, могут менять пространственную организацию нуклеиновых кислот. Полный или частичный разрыв водородных связей, ведущий к раскручиванию полинуклеотидных цепей нуклеиновой кислоты и их последующему разделению, называют денатурацией.
Липиды