

3.2. Клітинний центр. Центріолі
Клітинний центр – немембранна органела, розташована у тваринних клітинах поблизу ядра, у рослинних клітинах вона відсутня. Довжина клітинного центру складає близько 0,2-0,3 мкм, а діаметр – 0,1-0,15 мкм. Клітинний центр утворений двома центріолями, які лежать у взаємно перпендикулярних площинах, і променистою сферою з мікротрубочок. Клітинний центр бере участь в утворенні війок та джгутиків, у процесах збирання мікротрубочок та розподілі спадкового матеріалу клітини.
Центріолі – це щільні тільця. Центріолі(рис. 3.11) мають відносно постійне місце розташування в клітині: вони займають її геометричний центр, але іноді в процесі розвитку можуть переміщатися ближче до периферичних ділянок. Кожна центріоль утворена дев'ятьма групами мікротрубочок, зібраних по три – триплетами. Функція центріолі полягає в утворенні веретена поділу під час розмноження клітин. Крім того вони беруть участь в утворенні війок та джгутиків. [5]
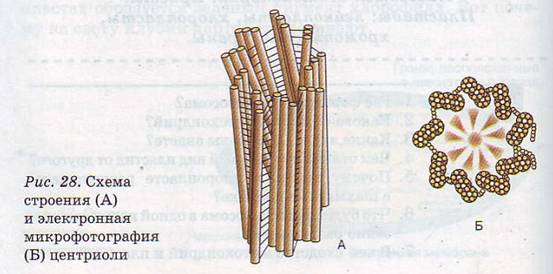
Рис. 3.11 Схема будови(А) і електронне фото(Б) центріолі
4. Молекулярні механізми руху
4.1. Прогрес в розумінні механізму руху клітин
Пересування клітин здійснюється за рахунок протрузії ведучого краю клітинної мембрани, координованого з переміщенням тіла клітини, і залежить від механізму роботи цитоплазматичних структур, здатних реагувати на сигнали від зовнішнього світу. Протрузія, як тепер стало зрозуміло, є наслідком полімеризації актину і було показано що GTP-ази сімейства Rho є проміжними ланками для передачі сигналів від мембранних рецепторів. Однак основна прогалина в розумінні регульованої рухливості заключалась у нерозумінні зв'язку між сигнальними шляхами і власне механізмом рухливості. Структурні, біохімічні та генетичні дослідження допомогли ідентифікувати деякі зв'язки, які були відсутні, і дозволили отримати обґрунтовану робочу модель, що описує шляхи поширення і механізми передачі і декодування сигналів, керуючих пересуванням клітин. [8]
4.2. Рух клітин і адгезійна взаємодія
Переміщення клітин визначається перебудовою адгезії до поверхні, на якій відбувається рух. Адгезійні локуси фомуються у результаті сигнальних взаємодій між позаклітинним матриксом і актиновим цитоскелетом всередині клітини, і вони асоціюються зі специфічними структурами актинових філаментів. Дослідження з використанням методів інтерференційної відбивної мікроскопії (IRM) вперше дозволили показати, що клітини не прилипають рівномірно до поверхні, але прикріплюються до неї тільки у окремих локусах (точках), найбільші з яких були названі фокальними контактами або точками фокальній адгезії. З інтерференційних картинок на кадрах IRM було встановлено, що відстані від клітин до субстрату у фокальних контактах знаходяться у діапазоні 10-15 мікрон. Такі ж дослідження виявили загальну нерухомість фокальних контактів щодо субстрату, що узгоджується з функцією адгезії. Адгезивна природа цих локусів була підтверджена у експериментах, де клітини були механічно видалені з поверхні на якій вони вирощувалися: після такої обробки у місцях фокальних контактів залишалися залишки, ізольовані і все ще прикріплені до субстрату Розрізняють дві широкі категорії адгезійних структур (локусів): 1) Фокальні комплекси, асоційовані з ламелоподіями і філоподіями, які підтримують клітинну протрузію і ретракцію на передньому фронті. 2) фокальні точки адгезії на кінцях пучків стресових фібрил, які відповідальні за більш стабільну адгезію. [14]