

§2 Высшая форма регуляции активности: Рассудочное поведение
Вопрос – «Может пи машина мыслить?»
Ответ – «Также нет!»
Дискуссия о Машине. 1956 г.
Разница между немецкими и американскими
лабораторными крысами состоит в том, что
в Америке эти животные снуют и суетятся,
обучаясь методом проб и ошибок, а в Германии
крысы сначала сидят и обдумывают свои действия...
Научный фольклор
Рассудочное поведение с точки зрения зоопсихолога
Попробуем посмотреть глазами биолога – что представляет собой «рассудочное» или «эвристическое» поведение? Что отличает его от рефлекса и инстинкта, что позволяет рассматривать его в качестве особого типа адаптивной активности и, наконец, свойственно ли оно лишь человеку?
Крупнейшие психологи и физиологи XIX и XX века вставали в тупик перед этой проблемой. Так, Вильгельм Вундт, основатель экспериментальной психологии, полагал, что лишь элементарные процессы ощущения, восприятия, внимания и памяти, которые подчиняются элементарным естественным законам, только и доступны для научного (физиологического) объяснения. Духовные же явления, которые есть категории человеческого духа, объяснить нельзя. Позднее Чарльз Шерринготон, который наряду с И. П. Павловым был основоположником теории рефлексов, в книге «Психика и мозг» выдвигал положение, что физиолог принципиально не может объяснить духовный мир человека, поскольку мир отвлечённых категорий существует вне человеческого мозга. Впрочем, мы уже знаем и о существовании других, «редукционистских» подходов к проблеме рассудочного поведения и мышления, но понимание трудности проблемы необходимо постоянно иметь в виду. И всё-таки что-то отличает поведение позвоночных животных и человека. Приведем пример, который открывает возможности к поиску ответа на данный вопрос – это факт из сравнительной психологии, известный как опыт Бойтендайка. Наблюдения проводились над рядом животных, принадлежащих к различным видам: над птицами, собаками, обезьянами.
П еред
животным ставили несколько рядов баночек
(рис. 7). Прямо на глазах животного в
первую банку первого ряда помещалась
приманка. Естественно, что животное
бежало к этой банке, перевертывало ее
и брало приманку. В следующий раз приманка
помещалась под вторую баночку, и если
только животное не видело эту приманку,
помещённую под новой баночкой, оно
бежало к прежней банке, и лишь затем, не
найдя приманки, бежало ко второй, где и
получало приманку. Так повторялось
несколько раз, причём каждый раз приманка
помещалась под следующую
баночку.
Оказалось, что ни одно животное не может
разрешить правильно эту задачу и сразу
бежать к следующей баночке, то есть оно
не может «схватить» принцип, что приманка
перемещается в каждую следующую баночку
ряда. В поведении животного доминируют
следы прежнего наглядного опыта и
отвлечённый принцип «следующий» не
формируется. В отличие от этого, маленький
ребёнок, примерно около 3,5–4 лет, легко
«схватывает» принцип «следующий» и уже
через несколько опытов тянется к той
баночке, которая раньше
никогда
не
подкреплялась, но
которая соответствует принципу
перемещения приманки на следующее
место.
еред
животным ставили несколько рядов баночек
(рис. 7). Прямо на глазах животного в
первую банку первого ряда помещалась
приманка. Естественно, что животное
бежало к этой банке, перевертывало ее
и брало приманку. В следующий раз приманка
помещалась под вторую баночку, и если
только животное не видело эту приманку,
помещённую под новой баночкой, оно
бежало к прежней банке, и лишь затем, не
найдя приманки, бежало ко второй, где и
получало приманку. Так повторялось
несколько раз, причём каждый раз приманка
помещалась под следующую
баночку.
Оказалось, что ни одно животное не может
разрешить правильно эту задачу и сразу
бежать к следующей баночке, то есть оно
не может «схватить» принцип, что приманка
перемещается в каждую следующую баночку
ряда. В поведении животного доминируют
следы прежнего наглядного опыта и
отвлечённый принцип «следующий» не
формируется. В отличие от этого, маленький
ребёнок, примерно около 3,5–4 лет, легко
«схватывает» принцип «следующий» и уже
через несколько опытов тянется к той
баночке, которая раньше
никогда
не
подкреплялась, но
которая соответствует принципу
перемещения приманки на следующее
место.
П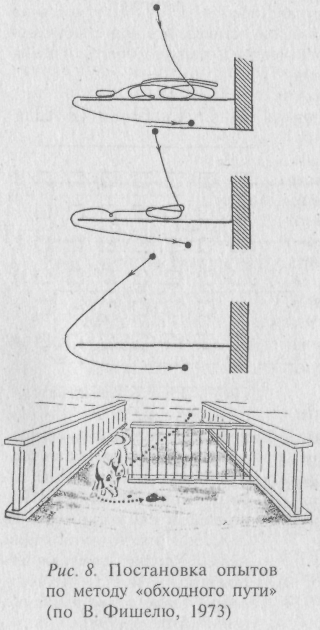 ри
каких-то обстоятельствах описанный
опыт, отвечающий на
вопрос
–
чего
они не
могут, можно
считать исчерпывающим для объяснения
различий между рациональным мышлением
человека и целесообразным поведением
животных. Для зоопсихолога важны ответы
и на другие вопросы: какие они, что у нас
общего, в чём именно и почему мы не такие?
ри
каких-то обстоятельствах описанный
опыт, отвечающий на
вопрос
–
чего
они не
могут, можно
считать исчерпывающим для объяснения
различий между рациональным мышлением
человека и целесообразным поведением
животных. Для зоопсихолога важны ответы
и на другие вопросы: какие они, что у нас
общего, в чём именно и почему мы не такие?
Рассматривая проблему с названных позиции, можно обсудить ряд опытов и наблюдений, посвящённых исследованию «элмементарной рассудочной деятельности» животных. Начинали её изучение с простого - перед голодным животным ставили кормушку, отделённую прозрачным препятствием (решеткой) и наблюдали, со скольки предъявлений оно научается безошибочно обходить препятствие. Опыт получил название «метода обходного пути» (рис. 8). К числу более сложных исследований следует отнести работы отечественного нейрофизиолога Л. В. Крушинского, выполненные в 70–80 годы. Эксперименты проводились с многими видами животных и суть их такова: устанавливается большая непрозрачная ширма с вертикальной щелью посредине, По одну сторону ширмы помещается испытуемое животное, по другую – две одинаковые кормушки, одна из которых содержит излюбленный данным видом корм, а вторая - пуста. Животное имеет доступ к корму через щель и поедает его из кормушки в течение нескольких секунд. Сразу вслед за этим обе кормушки начинают двигаться в разные стороны. Ширина щели позволяет видеть начальное направление движения кормушек. Животное правильно решает задачу в том случае, если оно передвигается вслед за невидимым через ширму кормом и поджидает его появления у дальнего края ширмы (рис. 9-1). По существу, этот эксперимент даёт возможность судить о том, могут ли животные строить простые планы своего будущего поведения, исходя из прогноза внешних явлений.
Животные разных видов в разной мере способны к построению таких планов. Самая древняя группа позвоночных среди исследованных – рыбы – с равной частотой следуют к полной кормушке, как и к пустой. У черепах и ящериц количество правильных решений значительно выше, чем неправильных. Среди птиц голуби подобны рыбам и совершенно не способны решать подобные задачи, а вот куры ведут себя много разумнее голубей, но уступают как ящерицам, гак и черепахам. Среди млекопитающих хуже всего успевают в решении задач названного типа кролики (примерно на уровне голубей). Значительно более способными оказались крысы, собаки, лисицы и волки. Волки, несомненно, наиболее способные в этом ряду прогнозисты, и ни один волк вообще ни разу не ошибся в выборе направления движения кормушки с едой.
В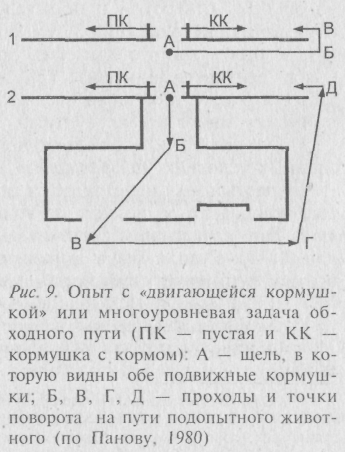 след
за этим исследователи решили усложнить
опыт – задачу. К ширме, за которой
двигались кормушки, с той стороны, где
находились животные, был пристроен
простой лабиринт (рис. 9-2). Теперь животное
не просто «исчисляло» движение нужной,
но невидимой кормушки, а вынуждено
менять направление движения, сначала
удаляясь от ширмы, затем, меняя направление
ещё раз, чтобы обогнуть преграду, минуя
ложные входы в лабиринте, только тогда
выходя к месту, где появляется корм.
Оказалось, что в данном случае решение
задачи оказалось непосильным не только
для птиц, но и для крыс. Только кошки,
собаки, лисицы и волки смогли решить
такую многоступенчатую задачу. Причём,
домашние кошки и собаки значительно
отставали от лис и волков.
след
за этим исследователи решили усложнить
опыт – задачу. К ширме, за которой
двигались кормушки, с той стороны, где
находились животные, был пристроен
простой лабиринт (рис. 9-2). Теперь животное
не просто «исчисляло» движение нужной,
но невидимой кормушки, а вынуждено
менять направление движения, сначала
удаляясь от ширмы, затем, меняя направление
ещё раз, чтобы обогнуть преграду, минуя
ложные входы в лабиринте, только тогда
выходя к месту, где появляется корм.
Оказалось, что в данном случае решение
задачи оказалось непосильным не только
для птиц, но и для крыс. Только кошки,
собаки, лисицы и волки смогли решить
такую многоступенчатую задачу. Причём,
домашние кошки и собаки значительно
отставали от лис и волков.
Конечно, на способность построения многоступенчатых планов были испытаны и высшие приматы. Шимпанзе строят одноступенчатые планы не задумываясь. Если перед клеткой лежит плод, до которого они не могут дотянуться, то будет использована палка, если она достаточно длинна. Двуступенчатый план строится с трудом. Так, немецкий зоопсихолог Вольфганг Кёлер положил в клетку шимпанзе Султану две палки, которые можно было вставить друг в друга, и положил банан так, чтобы его нельзя было достать ни одной из палок по отдельности, но лишь удлинённым «орудием». Задача была решена Султаном лишь через час. Правда, дочке Келера, которой в то время было менее трёх лет, на решение аналогичной задачи потребовалось несколько часов.
Трёхступенчатые задачи решаются шимпанзе с огромным трудом, особенно если в решении допускаются какие-то альтернативы. В опытах отечественного физиолога Э. Г. Вацуро, шимпанзе Рафаэль должен был соединить две палки – толстую с отверстием и тонкую (как и в описанном выше случае), но часто в ходе работы он просто забывал, ради чего всё это затеял. На следующий день, многократно собирая и разбирая свою конструкцию, Рафаэль в конце концов доставал плод составной палкой. Через несколько дней ему была предложена палка, которая помимо отверстия на конце имела ещё три боковых отверстия. Помня, что приманку можно достать составной палкой, Рафаэль повёл себя не самым разумным образом. Сначала он вставил тонкую палку в одно из боковых отверстий и попытался достать плод этой нелепой Г-образной конструкцией. Затем обезьяна поменяла её на Т-образную и опять тщетно пыталась дотянуться до плода. Когда после долгих мытарств шимпанзе пришел к верному решению, он опять забывал о конечной цели всех своих мучений и начинал доставать приманку много времени спустя.
Такие ситуации наблюдают нейрофизиологи в искусственно созданных условиях лабораторного эксперимента.
Существенно иначе видится поведение обезьян этологам, которые наблюдают животных на воле, в естественных для них условиях. Так, известная исследовательница поведения приматов Джейн ван Лавик-Гудолл была поражена, увидев впервые, как они добывают термитов, чтобы полакомиться ими:
На восьмой день моей бессменной вахты к термитнику снова подошел Дэвид Седобородый, на этот раз в сопровождении Голиафа. Они трудились у гнезда почти два часа. За это время я смогла увидеть много любопытных деталей: как они расковыривают свежезаделанные отверстия большим и указательным пальцами; как откусывают конец травинки, если он обломался, или же используют противоположный, целый конец; как отбрасывают одни орудия и находят другие. Голиаф однажды отошёл от термитника в поисках подходящего орудия метров на пятнадцать. Нередко оба самца срывали сразу три-четыре стебелька и клали рядом с термитником, используя их по мере надобности Но. пожалуй, самым интересным было то, как они подбирали небольшие веточки или плети лианы и, пропустив сквозь сжатый кулак, очищали их от листьев, делая пригодными к употреблению. Это можно считать первым документированным подтверждением того, что дикое животное не просто использует предмет в качестве орудия, но действительно изменяет его в соответствии со своими нуждами, демонстрируя тем самым зачатки изготовления орудий.
Таким образом, выяснилось, что многие высшие животные способны к элементарной рассудочной деятельности. Они могут осуществлять многоходовое целеполагание своих действий. Вместе с тем, они не в состоянии осуществить произвольно (не инстинктивную) сконструированную программу, на которую способен пятилетний ребенок. Почему? Вероятно потому, что оно не в состоянии: 1) построить её в уме; 2) удержать её в памяти в сокращённой и упрощённой, по сравнению с реальностью, целесообразной последовательности; 3) вспомнить и осуществить как дискретную последовательность действий.
Каковы же предпосылки в организации мозга, благодаря которым возникло столь сложное явление, как рассудочное поведение? Современная наука предлагает несколько положений, обобщающих основные данные нейрофизиологии, нейроанатомии и генетики, объединённых в гипотезу о механизмах рассудочной деятельности.
Структурные предпосылки рассудочного поведения
Первое положение. Большой объём мозга создаёт больше возможностей для упорядоченности его функций.
По сути – это общий принцип системной организации, полагающий, что в системах с большим количеством элементов создаётся больше предпосылок для качественного наращивания организации. В приложении к биологии он был впервые сформулирован физиком Э. Шредингером. Им было высказано предположение, что при восприятии мозгом физической организации внешней среды процессы, происходящие в мозге и регистрирующие «внешний порядок», также должны обладать чертами упорядоченности. По этой причине - чем большее число нейронов включается в осуществление происходящего в мозге процесса отражения мира, тем больше вероятность упорядоченного течения процесса. Биологически адекватное адаптивное поведение животного, которое обусловлено наиболее полным улавливанием закономерностей, связывающим предметы и явления окружающей среды, становится тем более продвинутым и гибким, чем выше упорядоченность работы мозга. Чем сложнее структура решаемых животным задач, тем большее число дискретных единиц, нейронов, должно быть вовлечено в её решение. Наиболее полное и точное улавливание многообразных закономерностей окружающего мира может быть осуществлено мозгом, состоящим из большего числа нейронов, а значит, и большего объёма.
Второе положение. Для более детального восприятия среды, которое необходимо для решения логических задач, необходимо чтобы отдельные нейроны объединялись в множество функциональных констелляций. Морфологическим аппаратом объединения нейронов являются известные нам синапсы.
Сравнительные морфологические исследования, выполненные различными исследователями на разных группах животных, показали, что существует вполне ясная зависимость между степенью ветвления дендритов в нейронах головного мозга, количеством синапсов (шипиков) на этих дендритах и способностью этих животных к решению экстраполяционных задач. В общем виде, очевидно, можно говорить, чем многочисленнее синаптические контакты нейронов, тем большая вероятность, что животное обладает более развитой рассудочной деятельностью.
Третье положение. В сером веществе коры мозга, в так называемых «экранных» его структурах, любые операции выполняются «дробно», локально.
Это обусловлено тем, что нейронные ансамбли коры головного мозга и мозжечка структурно оформлены в особого рода вертикальные колонки, пронизывающие всю толщу коры и называемые в настоящее время функциональными «модулями» головного мозга (рис. 10).
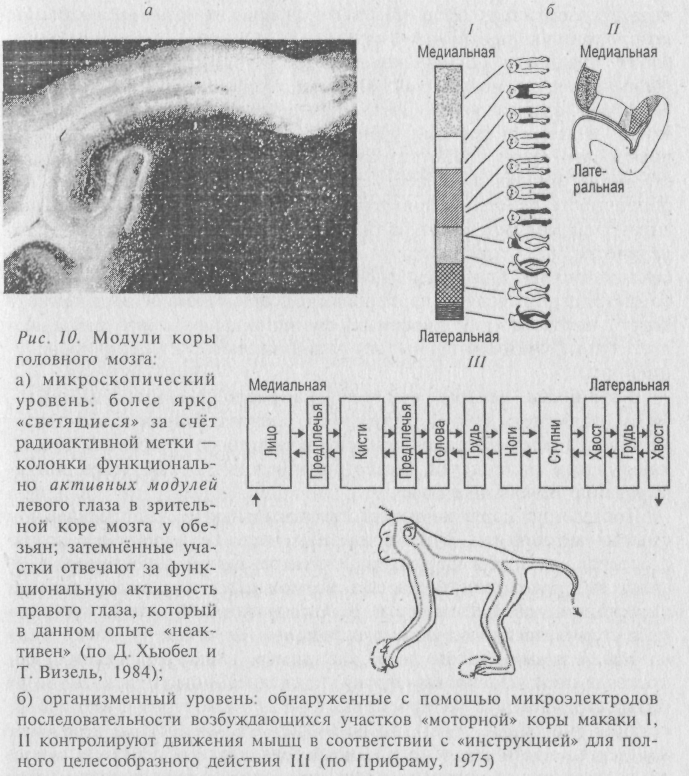
В состав таких колонок, представляющих собой вертикальные цилиндры диаметром около 30 мкм, входит около 110 нейронов. По некоторым подсчетам, только в неокортексе человека содержится около 600 миллионов «микроколонок», которые объединяются в более крупные рабочие комплексы, насчитывающие 1–3 миллиона нейронов. Колонки чувствительны к воздействию сигналов только определённого типа (здесь есть входное тестирующее устройство); они умножают распознанные сигналы, а затем распределяют их через систему горизонтальных нейронов лишь к вполне определённым, близким или далеким модулям-соседям. По мнению нейробиологов В. Маунткасла, Я. Сентаготаи, Дж. Экклса, модули являются функциональными единицами анализатора признаков. Еще в 1970 году физиолог Спинелли составил компьютерную программу (названную «Оккам»), моделирующую работу анализатора признаков и распознающую, реагирующую на волны определённой формы Особенно хорошо изучены колонки-детекторы в зрительной коре кошек сначала Д. Хьюбелом и Т. Визелем, а затем и другими учёными. Такие модули способны анализировать такие свойства раздражителя, как положение в пространстве, направление перемещения, степень его новизны и периодичность действия (появление и исчезновение). Конфигурации ансамблей модулей могут быть более жёстко детерминированы генетически, чем конфигурации нейронных ансамблей (а тем более синаптических). Благодаря такому явлению и стало возможным анатомо-топографическое картирование коры мозга и выделение в коре у человека (и у животных) функциональных зон, различающихся по функциям. Зоны эти в значительной степени видоспецифичны.
В сущности, центры речи человека, расположенные в доминантном (обычно в левом) полушарии, являются врождённо-детерминированными упорядоченными скоплениями нейронов, образующих ансамбли модулей, без которых невозможно обучение и оперирование речевыми сигналами.
Построение коры из множества локальных и, в определённом смысле, автономных функциональных модулей, вероятно и создает предпосылки для формирования «отраженной» дискретной картины действительности, то есть возможности для выделения неких свойств действительности, её «признаков». Вместе с этим, появляются и предпосылки к произвольным операциям с дискретными «элементами» такого мира (например, к экстраполяции и сопоставлению). Поскольку принцип дискретности проекции мира распространяется и на проекцию в мозге некоторых собственных органов (моторные зоны), то возникает возможность к произвольной деятельности животного. Дискретная картина действительности, запомненная животным, позволяет легко её декодировать, когда будет необходимо решить задачу с привлечением прошлого опыта.
Четвертое положение. Связано с формированием двух новых, по-видимому, взаимосвязанных свойств мозга, которые делают его по-настоящему способным не только к анализу, но и к синтезу. 1) Появлением особого механизма «сознания», механизма, который способствует оперативному отбору тех констелляций нейронов, которые необходимы для решения «логических» задач, имеющих биологическую значимость. 2) Способности к оценке значимости решаемой задачи, осуществляемой эмоциями.
Ясно, что у психологов вызовет недоумение слово «сознание», отнесённое к животным. Нейрофизиологи употребляют этот термин, сознательно редуцируя его содержание:
Под сознанием мы понимаем функцию восприятия мозгом текущих событий окружающей действительности, на основе которой происходит синтез всего пережитого индивидуального опыта. (Этим достигается осознание окружающей действительности). Говоря о сознании, мы имеем в виду, конечно, не общественное сознание, которое присуще только человеку. Под сознанием мы понимаем лишь восприятие окружающей действительности в текущий момент и возможность его сравнения с восприятиями прошлого. Это есть то сознание, которое теряет человек и животное, например, во время наркоза, и которое возвращается после его прекращения.
Любопытно, что сознание (в смысле conscious) связано отнюдь не только с неокортексом, главным атрибутом головного мозга человека. Так, при проведении многочисленных операций на мозге людей, находящихся во время операции в полном сознании, выяснилось, что удаление различных отделов коры больших полушарий никогда не приводит к потере сознания пациента. Однако сознание теряется в тех случаях, когда блокируется, функциональная активность более древней части – промежуточного мозга. Хотя никто не считает таламус или какие-то его структуры вместилищем сознания, тем не менее, У. Пенфилд и Г. Джаспер выделили комплекс «таламус – кора больших полушарий» в качестве особой центрэнцефалической системы. По мнению У. Пенфилда, многие интегративные процессы, лежащие в основе произвольных движений и сознания, осуществляются при участии центрэнцефалической системы.
Взаимообусловленность рассудочного поведения и эмоций
Ещё в 1937 году Дж. Пайпец высказал предположение, что передние ядра таламуса, гипоталамус, поясная извилина коры мозга и гиппокамп образуют единый механизм, работа которого и является основой для возникновения эмоций. Совокупность этих структур долгое время так и называлась – круг Пайпеца (рис. 11). Позднее, учитывая, что поясная извилина как бы окаймляет основание переднего мозга, было предложено другое название – лимбическая система. Считается, что источником возбуждения для этой системы является гипоталамус. В ядрах гипоталамуса, по-видимому, находят своё материальное представительство и многие инстинктивные программы животных и человека.
Профессор Л. В. Крушинский приводит опыт, который иллюстрирует роль эмоций в успехе решения экстраполяционных задач. Крысы лабораторных линий не в состоянии решать экстраполяционную задачу при использовании пищевого раздражителя в качестве стимула к её решению. Однако, если вместо этого используется прямое электрическое раздражение лимбической системы мозга, которое крыса осуществляет сама, через вживлённые электроды посредством нажатия на педаль, то она вполне успешно решает такие задачи. Они преследуют отодвигающуюся за непрозрачной ширмой педаль и прогнозируют направление движения педали, предвкушая вожделенное ими раздражение «центров удовольствия» после встречи с нею. Принципиальное значение описанных опытов заключается в том, что ясно показывает роль непосредственного раздражения структур мозга, связанных с эмоциями при выполнении элементарного рассудочного акта, а значит – и роль тех самых эмоциональных влиянии для выбора адекватной формы поведения.
Ф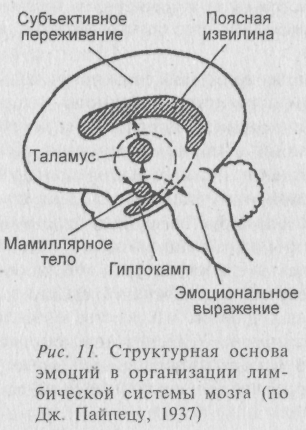 ундаментальное
значение открытия необходимости
эмоциональной поддержки для решения
задач, связанных с адаптивным поведением,
трудно переоценить. В условиях реальности,
построенной на неопределённостях, да
ещё при наличии многоступенчатых
разветвлённых планов, где очень часто
приходится совершать выбор
на
основе неполной информации, без мотиваций,
без влечений, задачи становятся просто
нерешаемыми.
ундаментальное
значение открытия необходимости
эмоциональной поддержки для решения
задач, связанных с адаптивным поведением,
трудно переоценить. В условиях реальности,
построенной на неопределённостях, да
ещё при наличии многоступенчатых
разветвлённых планов, где очень часто
приходится совершать выбор
на
основе неполной информации, без мотиваций,
без влечений, задачи становятся просто
нерешаемыми.
Таким образом, основы эволюции структур мозга, которые связаны с рассудочной деятельностью, определялись становлением трёх его морфо-функциональных комплексов:
а) комплекса «ствол мозга – передний отдел головного мозга». Видимо, это самый древний функциональный комплекс, и его характер весьма сходен у многих животных, а развитие выражено главным образом количественно;
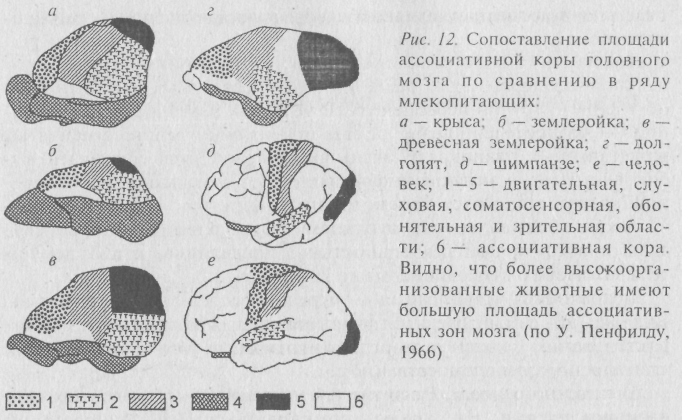
б) конечного мозга. Развитие его у позвоночных животных шло разными путями. Но, кроме упоминавшихся новоприобретений мозга за счёт этой зоны, а именно – увеличения объёма мозга и усложнения системы синаптических контактов, увеличивается площадь так называемых ассоциативных зон, или зон взаимного перекрывания специализированных областей сенсорных центров в коре. Относительное прогрессивное увеличение ассоциативных зон, к которым относится лобная область, по отношению к специализированным отделам коры (обонятельной, зрительной, слуховой и других) отчётливо видно в сравнительном ряду мозга млекопитающих (рис. 12);
в) лимбической системы. Её филогенез находился в тесной связи с развитием новой коры (рис. 13). Старая кора (архикортекс), первоначально функционально связанная с дистальной (обонятельной) рецепцией, оказалась у млекопитающих оттеснённой на медиальную сторону и вошла в состав лимбической системы. У высших млекопитающих лимбическая система достигает наивысшей дифференциации. А у человека и шимпанзе задняя лимбическая система фактически неотличима от других структур новой коры и, по-видимому, входит в её состав. Некоторые исследователи находят, что такая прогрессивная структура новой коры, как префронтальная область, может быть отнесена к структурам лимбической системы.
Наконец, надо иметь в виду, что основу индивидуального адаптивного поведения составляют процессы обучения и памяти, а также сложно устроенные и недостаточно изученные механизмы научения, запоминания и воспоминания (воспроизведения).
П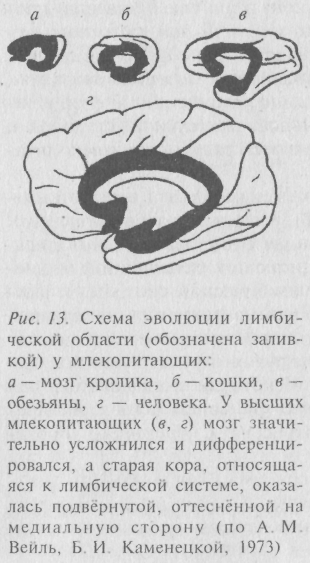 амять
генная и память мозга сходны в том, что
их главный «секрет» во внутренней
дискретности устройства,
а их эффективность обусловлена тем, что
какие-то части внешнего мира могут
«ложиться» в ячейки гой и другой памяти,
превращаясь в «знаки»
этого
мира, отражая те самые свойства. В целом
же создаются значимые
Образы
мира, которые
теперь могут быть выражены в «символах»,
что
в общем-то и есть образы действительности,
но получившие эмоциональную
оценку.
амять
генная и память мозга сходны в том, что
их главный «секрет» во внутренней
дискретности устройства,
а их эффективность обусловлена тем, что
какие-то части внешнего мира могут
«ложиться» в ячейки гой и другой памяти,
превращаясь в «знаки»
этого
мира, отражая те самые свойства. В целом
же создаются значимые
Образы
мира, которые
теперь могут быть выражены в «символах»,
что
в общем-то и есть образы действительности,
но получившие эмоциональную
оценку.
Вопрос о роли знаков и символов подробнее будет обсуждаться дальше в рамках проблемы коммуникаций (см. Глава 4) в мире животных. Заметим только, что именно дискретность экранных структур мозга (новой коры у млекопитающих, полосатого тела у птиц) способствовала появлению эффективных знаковых средств, совершенного социального поведения, рассудочного поведения и, наконец, самого рассудка.
Об асимметрии мозга и «двойственности» Разума
О неравноправии правой и левой половин тела человека было известно ещё Гиппократу. В те времена, когда не знали о связи головного мозга и душевных функций, считалось, что правая половина тела находится под контролем Меркурия – планеты «интеллекта», а левая – под контролем Марса, олицетворяющего импульсивность и гневливость. Сегодня бы сказали, что Меркурий владеет левым полушарием мозга, а Марс – правым. Знания о неравнозначности полушарий мозга у людей особенно интенсивно накапливались в течение последних ста лет. При этом очень долго исследования практически не проводились на животных, ибо это считалось бесперспективным. Ведь морфо-функциональная асимметричность предполагалась как отличительный признак человеческого мозга.
При этом анатомы утверждали, что полушария головного мозга человека отличаются незначительно. Их различия не больше, чем вариации формы мозга у разных людей. Но функциональные различия правого и левого полушарий очень велики как у людей, так и у животных. Самым очевидным проявлением латерализации мозга у человека является неравенство правой и левой руки. Это явление по-настоящему загадочно. До сих пор существует мнение, что асимметрия мозга тесно связана с высшими языковыми (речевыми) функциями. Именно с «лёгкой руки» авторитетного французского невропатолога и анатома XIX века Поля Брока, открывшего в левом полушарии головного мозга двигательные речевые зоны (зоны Брока), долго считалось, что асимметрия мозга присуща исключительно людям. И как раз она определяет наши уникальные языковые способности. Данное представление вызывалось тем, что межполушарных различий у животных, даже у ближайших к человеку обезьян, учёные не обнаруживали.
И всё-таки сначала удалось обнаружить, что многие виды животных (но в разной мере) проявляют латерализацию. В 70-е годы Р. Коллинз и Дж. Уоррен показали, что обезьяны, кошки и мыши предпочитают пользоваться одной и той же конечностью (правой или левой), если задача не требует подключения другой. Однако
распределение животных, предпочитающих пользоваться той или другой лапой, приблизительно составляет 50 : 50. То есть у животных предпочтение это случайное, оно может быть не очень выраженным, хотя факт латерализации здесь имеет место. В то время, как у людей 90% предпочитает правую руку и только 10% -левую.
Вскоре был сделан ряд замечательных открытий, указывающих на существовании видов, у которых одно полушарие доминирует над другим. В этом отношении особый интерес представляли исследования асимметрии мозга у птиц, опубликованные в 1966 году Ф. Ноттебом и его коллегами. Они продемонстрировали, что перерезка у взрослых зябликов и канареек левого подъязычного нерва, который управляет работой мышц левой половины гортани, до неузнаваемости изменяет их пение. Но перерезка правого подъязычного нерва, управляющего правой половиной гортани, влечёт минимальные изменения в характере их пения. Это были первые сведения о том, что мозг у птиц обладает выраженной функциональной (левой) асимметрией, что и было подтверждено позднее с помощью самых современных методов в многочисленных работах нейробиологов.
Более того, в последнее время признаётся, что асимметричное функционирование больших полушарий головного мозга у представителей животного мира является фундаментальной (хотя и неразгаданной) закономерностью его деятельности.
Современные биологи научились выявлять скрытую (латентную) асимметрию мозга у животных. Так, в исследованиях М. Петерсена с коллегами (1978), проведённых на японских макаках, обнаружена левосторонняя асимметричность их полушарий по восприятию звуковых сигналов. Обезьян обучали различать два типа звуков, издаваемых особями их вида и определённым образом реагировать на них. Звуки предварительно записывались на магнитофон и предъявлялись в правое или левое ухо животного в случайном порядке. Обнаружилось, что макаки более точно осуществляли требуемые реакции при трансляции сигнала на правое ухо.
По утверждению В.Л. Бианки, имеются многочисленные данные о двигательно-пространственных асимметриях даже у беспозвоночных животных: планарий, мучнистого хрущака, речных раков, муравьев и других. Бианки предложил синтетическую модель межполушарных отклонений, опирающуюся на три принципа деятельности мозга: асимметрии, доминантности и комплементарности. По его мнению, у животных существуют два основных способа описания мира, которые специфично реализуются в полушариях мозга – это индукция и дедукция. При индуктивной обработке анализ предшествует синтезу, а при дедуктивной – наоборот, синтез предшествует анализу. Не следует думать, что каждое из полушарий строго специализировано либо на синтезе, либо на анализе. Нет, они оба участвуют в названных процессах, но при реализации разных способов обработки информации доминирует одно из них.
В левом полушарии преобладают:
– индуктивная обработка;
- восприятие абстрактных признаков;
– последовательная обработка;
- аналитическое восприятие;
– восприятие времени. В правом полушарии:
– дедуктивная обработка;
- восприятие конкретных признаков;
– одновременная обработка;
– примат синтетического (целостного) восприятия;
- восприятие пространства.
По-прежнему мало ясности в вопросе о наследуемости функциональной асимметрии полушарий, а следовательно, и нет объяснения такой видовой особенности человека, как выраженная праворукость (доминирование левого полушария). В 1969 году генетик Р. Коллинз провел опыты по скрещиванию мышей с одинаково предпочитаемой лапой. Он трижды повторил близкородственное (инбредное) скрещивание и проверил соотношение лево- и праволапого потомства в последнем, третьем, поколении. Результаты свидетельствовали, что генетического контроля за латеральными предпочтениями у мышей нет. В потомстве исключительно леволапых, равно как и праволапых особей, повторялось исходное (как и до опыта) распределение 50 : 50.
Глава III
ИНДИВИДУАЛЬНАЯ ПАМЯТЬ И НАУЧЕНИЕ
Введение. Условия обогащения поведения
§ 1 ПРИВЫКАНИЕ И СЕНСИТАЦИЯ – ПРОСТЕЙШИЕ ФОРМЫ НАУЧЕНИЯ
§ 2 ПАМЯТЬ И НАУЧЕНИЕ КАК ФУНКЦИИ ГОЛОВНОГО МОЗГА
– Мозг как инструмент запоминания и научения
– Пошаговое научение посредством механизма «стимул – реакция»
§ 3 ФОРМЫ НАУЧЕНИЯ, НЕ СВЯЗАННЫЕ С ПОДКРЕПЛЕНИЕМ
– Латентное научение
– Условнорефлекторное избегание пищи
– Имитационное научение
– Эвристическое научение
– Исследование отсроченных реакций
– Импринтинг
§ 4 СТРУКТУРНО-ФУНКЦИОНАЛЬНЫЕ ОСНОВЫ ПАМЯТИ И НАУЧЕНИЯ
– Виды памяти
§ 5 ПОИСКИ МЕХАНИЗМОВ ПАМЯТИ
– Истоки: малые системы нейронов
– От природных моделей к культуре нейронов: история с «обучающимся блюдцем •>
– Локализация памяти в головном мозге (вид Homo sapiens)
– Долговременная потенциация в гиппокампе
– Гиппокамп как карта активного поведения
– Нейродинамика памяти (или истории с «куриным мозгами»)
§ 6 ЗАВИСИМОСТЬ МОЗГА ОТ СВОЙСТВ ВНЕШНЕЙ СРЕДЫ
- Влияние возрастных факторов
– Пренатальный период
- Период выкармливания
– Периоды после вскармливания
– Влияние средовых факторов

Глава III