

1 Методы исследования функций центральной нервной системы : Функции нервной системы изучают с использованием традиционных классических для общей физиологии методов и специальных методических подходов, призванных выявить специфические функции нервных образований, выполняющих роль главной управляющей и информационной системы в организме. В соответствии с двумя принципиально различными методическими подходами к изучению физиологических функций организма различают методы экспериментальной и теоретической нейрофизиологии. К числу экспериментальных методов классической физиологии относятся приемы, направленные на активацию, или стимуляцию, подавление, или угнетение, функции данного нервного образования. Способы активирования изучаемого органа сводятся к раздражению его адекватными (или неадекватными) стимулами. Адекватное раздражение достигается специфическим раздражением соответствующих рецептивных входов рефлексов либо электрическим раздражением проводникового или центрального отдела рефлекторной дуги, имитирующим нервные импульсы. Среди неадекватных стимулов наиболее распространенными являются раздражение различными химическими веществами и градуируемое раздражение электрическим током. Классический условнорефлекторный метод изучения функции коры большого мозга в современной нейрофизиологии успешно применяется в комплексном анализе механизмов обучения, становления и развития адаптивного поведения в сочетании с методами электроэнцефалографии, электронейронографии, нейро- и гистохимии, психофизиологии, способствуя более полному представлению физиологической сущности протекающих в мозге процессов.
Регуляция физиологических функций происходит за счет трех регуляторных систем – нервной, эндокринной и иммунной. В целом выделяют два ведущих пути регуляции – нервный и гуморальный. Каким бы не был путь регуляции механизм его в целом един. Схематически регуляторную систему можно представить в виде трех составляющих ее компонентов: 1. Собственно регуляторная система, например, центральная нервная система. 2. Объект регуляции, им может быть любой исполнительный орган, например, сердце. 3. Измерительное устройство – это рецепторы, которые воспринимают информацию и передают ее в ЦНС, изменяя тем самым ее функционирование. Целью любого пути регуляции является достижение конечного эффекта в виде стабильного гомеостаза, т.е. в виде постоянства внутренней среды
Нейрогуморальная регуляция физиологических функций. Жизнедеятельность каждого организма должна находиться в строгом соответствии с условиями окружающей среды. Для этого каждое существо должно воспринимать сигналы внешней среды (свет, звук, температуру, давление и др.), усваивать, обрабатывать их и правильно на них реагировать. В этом случае весь организм должен выступать как единое целое, органы и системы органов которого работают согласованно, упорядоченно.
Такую согласованность, упорядоченность действий в организме человека выполняют два механизма: нервный и гуморальный. Их действием и влиянием на органы, системы органов осуществляется регуляция всех процессов жизнедеятельности организма, обеспечивается его целостность.
Нервная регуляция — регуляция жизнедеятельности организма с помощью нервной системы. Гуморальная регуляция осуществляется с помощью химических веществ через жидкие среды организма (кровь, лимфу, межклеточную жидкость).
Нервная и гуморальная регуляции функций организма взаимосвязаны. На функциональное состояние нервной системы оказывают влияние активные химические вещества, циркулирующие в крови, например гормоны (от греческого «гормао» — побуждать). Образование гормонов железами внутренней секреции и выделение их в кровь осуществляется под контролирующим влиянием нервной системы. В связи с этим следует рассматривать и говорить не о раздельном влиянии нервной системы и гуморальных факторов, а о едином механизме нейрогуморальной регуляции функций организма.
ажнейшее свойство организма — саморегуляция физиологических функций, которая автоматически поддерживает относительное постоянство внутренней среды организма — гомеостаз (от греческого «гомоис» — тот же самый и «стасис» — состояние), что является необходимым условием существования. Саморегуляция возможна потому, что имеются обратные связи между регулируемым процессом и регулирующей системой, когда информация о конечном результате поступает в центральную нервную систему.Нервная система представляет собой совокупность структур, которые регулируют работу отдельных органов и систем, осуществляют взаимосвязь отдельных органов между собой и всего организма с внешней средой.
2. Нейрон (от др.-греч. νεῦρον — волокно, нерв) — это структурно-функциональная единица нервной системы. Эта клетка имеет сложное строение, высоко специализирована и по структуре содержит ядро, тело клетки и отростки. В организме человека насчитывается более ста миллиардов нейронов. Нейроны различаются по форме, числу отростков и функциям. В зависимости от функции выделяют чувствительные, эффекторные(двигательные, секреторные) и вставочные. Чувствительные нейроны воспринимают раздражения, преобразуют их в нервные импульсы и передают в мозг. Эффекторные (от лат. эффектус - действие) - вырабатывают и посылают команды к рабочим органам. Вставочные - осуществляют связь между чувствительными и двигательными нейронами, учавствуют в обработке информации и выработке команд.
Безаксонные нейроны — небольшие клетки, сгруппированы вблизи спинного мозга в межпозвоночных ганглиях, не имеющие анатомических признаков разделения отростков на дендриты и аксоны. Все отростки у клетки очень похожи. Функциональное назначение безаксонных нейронов слабо изучено.
Униполярные нейроны — нейроны с одним отростком, присутствуют, например в сенсорном ядре тройничного нерва в среднем мозге.
Биполярные нейроны — нейроны, имеющие один аксон и один дендрит, расположенные в специализированных сенсорных органах — сетчатке глаза, обонятельном эпителии и луковице, слуховом и вестибулярном ганглиях.
Мультиполярные нейроны — нейроны с одним аксоном и несколькими дендритами. Данный вид нервных клеток преобладает в центральной нервной системе.
Псевдоуниполярные нейроны — являются уникальными в своём роде. От тела отходит один отросток, который сразу же Т-образно делится. Весь этот единый тракт покрыт миелиновой оболочкой и структурно представляет собой аксон, хотя по одной из ветвей возбуждение идёт не от, а к телу нейрона. Структурно дендритами являются разветвления на конце этого (периферического) отростка. Триггерной зоной является начало этого разветвления (то есть находится вне тела клетки). Такие нейроны встречаются в спинальных ганглиях.
Афферентные нейроны (чувствительный, сенсорный или рецепторный). К нейронам данного типа относятся первичные клетки органов чувств и псевдоуниполярные клетки, у которых дендриты имеют свободные окончания.
Эфферентные нейроны (эффекторный, двигательный или моторный). К нейронам данного типа относятся конечные нейроны — ультиматные и предпоследние — не ультиматные.
Ассоциативные нейроны (вставочные или интернейроны) — группа нейронов осуществляет связь между эфферентными и афферентными, их делят на интризитные, комиссуральные и проекционные.
Секреторные нейроны — нейроны, секретирующие высокоактивные вещества (нейрогормоны). У них хорошо развит комплекс Гольджи, аксон заканчивается аксовазальными синапсами.
3. Время рефлекса.Его составляющие компоненты и факторы влияющие на него.
Нейроны и пути прохождения нервных импульсов при рефлекторном акте образуют так называемую рефлекторную дугу: стимул — рецептор-аффектор — нейрон ЦНС — эффектор — реакция.
Рефлекс - это наиболее правильная, чаще всего встречающаяся реакция организма на внешние раздражители.
По типу образования: условные и безусловные
По видам рецепторов: экстероцептивные (кожные, зрительные, слуховые, обонятельные), интероцептивные (с рецепторов внутренних органов) и проприоцептивные (с рецепторов мышц, сухожилий, суставов)
По эффекторам: соматические, или двигательные (рефлексы скелетных мышц), например флексорные, экстензорные, локомоторные, статокинетические и др.; вегетативные внутренних органов — пищеварительные, сердечно-сосудистые, выделительные, секреторные и др.
По биологической значимости: оборонительные, или защитные, пищеварительные, половые, ориентировочные.
По степени сложности нейронной организации рефлекторных дуг различают моносинаптические, дуги которых состоят из афферентного и эфферентного нейронов (например, коленный), и полисинаптические, дуги которых содержат также 1 или несколько промежуточных нейронов и имеют 2 или несколько синаптических переключений (например, флексорный).
По характеру влияний на деятельность эффектора: возбудительные — вызывающими и усиливающими (облегчающими) его деятельность, тормозные — ослабляющими и подавляющими её (например, рефлекторное учащение сердечного ритма симпатическим нервом и урежение его или остановка сердца — блуждающим).
По анатомическому расположению центральной части рефлекторных дуг различают спинальные рефлексы и рефлексы головного мозга. В осуществлении спинальных рефлексов участвуют нейроны, расположенные в спинном мозге. Пример простейшего спинального рефлекса — отдергивание руки от острой булавки. Рефлексы головного мозга осуществляются при участии нейронов головного мозга. Среди них различают бульбарные, осуществляемые при участии нейронов продолговатого мозга; мезэнцефальные — с участием нейронов среднего мозга; кортикальные — с участием нейронов коры больших полушарий головного мозга.
Время рефлекса.
Время рефлекса - время от начала раздражения рецептора до появления ответной реакции организма. Время рефлекса складывается: - из времени возбуждения афферентных и эфферентных образований; - из времени проведения возбуждения по афферентным и эфферентным волокнам; - из времени переключения импульсации в центральных структурах мозга, участвующих в реализации рефлекса.
Время рефлекса зависит также от возбудимости НС в данный момент. При утомлении нервных центров время рефлекса увеличивается.
Последействие рефлекса.
Продолжительность рефлекса всегда больше, чем время раздражения. Это связано с тем, что возбуждение в нервных центрах циркулирует еще длительное время после действия раздражителя.
Возбуждение от одного нейрона к другому передается как по прямой цепи, так и по боковым замкнутым цепям вставочных нейронов.
4 Нервный центр — совокупность структур центральной нервной системы, координированная деятельность которых обеспечивает регуляцию отдельных функций организма или определенный рефлекторный акт. Представление о структурно-функциональной основе нервного центра обусловлено историей развития учения о локализации функций в центральной нервной системе. На смену старым теориям об узкой локализации, или эквипотенциальности, высших отделов головного мозга, в частности коры большого мозга, пришло современное представление о динамической локализации функций, основанное на признании существования четко локализованных ядерных структур нервных центров и менее определенных рассеянных элементов анализаторных систем мозга. При этом с цефализацией нервной системы растут удельный вес и значимость рассеянных элементов нервного центра, внося существенные различия в анатомических и физиологических границах нервного центра. В результате функциональный нервный центр может быть локализован в разных анатомических структурах. Например, дыхательный центр представлен нервными клетками, расположенными в спинном, продолговатом, промежуточном мозге, в коре большого мозга.
Нервные центры имеют ряд общих свойств, что во многом определяется структурой и функцией синаптических образований.
1. Односторонность проведения возбуждения. В рефлекторной дуге, включающей нервные центры, процесс возбуждения распространяется в одном направлении (от входа, афферентных путей к выходу, эфферентным путям).
2. Иррадиация возбуждения. Особенности структурной организации центральных нейронов, огромное число межнейронных соединений в нервных центрах существенно модифицируют (изменяют) направление распространения процесса возбуждения в зависимости от силы раздражителя и функционального состояния центральных нейронов. Значительное увеличение силы раздражителя приводит к расширению области вовлекаемых в процесс возбуждения центральных нейронов — иррадиации возбуждения.
3. Суммация возбуждения. В работе нервных центров значительное место занимают процессы пространственной и временной суммации возбуждения, основным нервным субстратом которой является постсинаптическая мембрана. Процесс пространственной суммации афферентных потоков возбуждения облегчается наличием на мембране нервной клетки сотен и тысяч синаптических контактов. Процессы временной суммации обусловлены суммацией ВПСП на постсинаптической мембране.
4. Наличие синаптической задержки. Время рефлекторной реакции зависит в основном от двух факторов: скорости движения возбуждения по нервным проводникам и времени распространения возбуждения с одной клетки на другую через синапс. При относительно высокой скорости распространения импульса по нервному проводнику основное время рефлекса приходится на синаптическую передачу возбуждения (синаптическая задержка). В нервных клетках высших животных и человека одна синаптическая задержка примерно равна 1 мс.
5. Высокая утомляемость. Длительное повторное раздражение рецептивного поля рефлекса приводит к ослаблению рефлекторной реакции вплоть до полного исчезновения, что называется утомлением. Этот процесс связан с деятельностью синапсов — в последних наступает истощение запасов медиатора, уменьшаются энергетические ресурсы, происходит адаптация постсинаптического рецептора к медиатору.
6. Тонус. Тонус, или наличие определенной фоновой активности нервного центра, определяется тем, что в покое в отсутствие специальных внешних раздражений определенное количество нервных клеток находится в состоянии постоянного возбуждения, генерирует фоновые импульсные потоки. Даже во сне в высших отделах мозга остается некоторое количество фоновоактивных нервных клеток, формирующих «сторожевые пункты» и определяющих некоторый тонус соответствующего нервного центра.
7. Пластичность. Функциональная возможность нервного центра существенно модифицировать картину осуществляемых рефлекторных реакций. Поэтому пластичность нервных центров тесно связана с изменением эффективности или направленности связей между нейронами.
8. Конвергенция. Нервные центры высших отделов мозга являются мощными коллекторами, собирающими разнородную афферентную информацию. Количественное соотношение периферических рецепторных и промежуточных центральных нейронов (10:1) предполагает значительную конвергенцию («сходимость») разномодальных сенсорных посылок на одни и те же центральные нейроны.
9. Интеграция в нервных центрах. Важные интегративные функции клеток нервных центров ассоциируются с интегративными процессами на системном уровне в плане образования функциональных объединений отдельных нервных центров в целях осуществления сложных координированных приспособительных целостных реакций организма (сложные адаптивные поведенческие акты).
10. Свойство доминанты. Доминантным называется временно господствующий в нервных центрах очаг (или доминантный центр) повышенной возбудимости в центральной нервной системе. По А.А.Ухтомскому, доминантный нервный очаг характеризуется такими свойствами, как повышенная возбудимость, стойкость и инертность возбуждения, способность к суммированию возбуждения.
11. Цефализация нервной системы. Основная тенденция в эволюционном развитии нервной системы проявляется в перемещении, сосредоточении функции регуляции и координации деятельности организма в головных отделах ЦНС. Этот процесс называется цефализацией управляющей функции нервной системы.
5. Одностороннее проведение возбуждения через синапсы. По нервным волокнам импульсы возбуждения способны распространяться в обе стороны от места раздражения. В центральной же нервной системе они распространяются обычно лишь в одном направлении - только с афферентных нейронов на эфферентные. Это означает, что в ЦНС импульсы передаются лишь с аксона одного нейрона на клеточное тело и дендриты других нейронов и не передаются с дендритов и с тела нервной клетки на подходящие к ним веточки аксона.
Указанная закономерность была впервые установлена в1823 году одновременно двумя исследователями - шотландцем И.Беллом и французским физиологом Ф.Мажанди - и получила название закона Белла-Мажанди, согласно которому афферентные волокна вступают в спинной мозг через задние корешки, а эфферентные волокна покидают спинной мозг через передние корешки.
Одностороннее проведение возбуждения в нервных центрах обусловлено строением синапсов: медиаторы выделяются только концевыми аппаратами аксонов и к медиаторам чувствительна только постсинаптическая мембрана синапса, на которой возникает потенциал действия (возбуждающий или тормозящий). Таким образом, возбуждение в синапсе распространяется от окончаний аксона через медиатор на постсинаптическую мембрану тела нервной клетки, дендрита или вставочного нейрона. В обратном направлении передача возбуждения возможна только в электрическом синапсе, в котором возбуждение от пресимпатической мембраны передается к постсинаптической электрическим путем.
Одностороннее проведение возбуждения - возбуждение передается с афферентного на эфферентный нейрон. Причина: клапанное свойство синапса.
Задержка проведения возбуждения: скорость проведения возбуждения в нервном центре на много ниже таковой по остальным компонентам рефлекторной дуги. Чем сложнее нервный центр, тем дольше проходит по нему нервный импульс. Причина: синаптическая задержка. Время проведения возбуждения через нервный центр - центральное время рефлекса.
6.Замедленное
проведение возбуждения.
В основе проведения нервных импульсов
по цепочке нейронов лежат два различных
механизма: электрический (проведение
потенциала действия — ПД — по нервным
волокнам) и химический (передача через
синапс с помощью медиатора). Первый
осуществляется с большой скоростью (до
100—140 м/сек), второй—в тысячу раз
медленнее. Замедление проведения связано
с затратой времени на процессы,
происходящие от момента прихода
пресинаптического импульса в синапс
до появления в постсинаптической
мембране возбуждающих или тормозных
потенциалов. Этот интервал называется
синаптической задержкой и составляет
в мотонейронах и большинстве других
клеток Центральной нервной системы
примерно 0,3 мсек. В вегетативной нервной
системе длительность синаптической
задержки больше — не менее 10 мсек. За
это время пресинаптический импульс
вызывает опорожнение синаптических
пузырьков, происходит диффузия медиатора
через синаптическую щель, увеличение
под его влиянием ионной проницаемости
постсинаптической мембраны и возникает
постсинаптический потенциал. После
этого требуется еще около 1,2 мсек. на
развитие возбуждающего постсинаптического
потенциала (ВПСП) до максимума, чтобы
возник (в случае достижения критического
уровня деполяризации) ПД. Таким образом,
весь процесс передачи нервного импульса
(от ПД одной клетки до ПД следующей
клетки) через один синапс занимает
примерно 1,5 мсек. При утомлении, охлаждении
и ряде других воздействий длительность
синаптической задержки возрастает.
Если же для осуществления какой-либо
реакции требуется участие большого
числа нейронов (многих сотен и даже
тысяч), то суммарная величина задержки
проведения по нервным центрам — так
называемое центральное время проведения
— может составить десятые д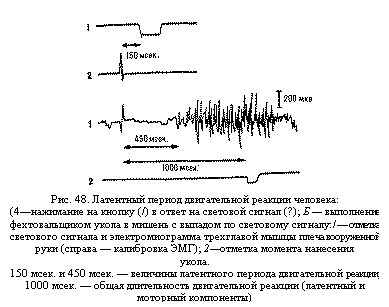 оли
секунды и даже целые секунды. Поскольку
проведение импульсов по нервным волокнам
от периферических рецепторов в нервные
центры и от нервных центров к исполнительным
органам занимает сравнительно небольшое
время, общее время от момента нанесения
внешнего раздражения до появления
ответной реакции организма (латентный
период рефлекса) определяется центральным
временем проведения. Величина латентного
периода рефлекса служит важным показателем
функционального состояния нервных
центров и широко используется в практике.
Латентный период двигательной реакции
измеряется от момента подачи сигнала
до момента начала движения (или до
появления электрической активности
соответствующей мышцы). При осуществлении
человеком простых движений в ответ на
внешние сигналы, например нажимание на
кнопку при световом сигнале (рис. 48, А),
длительность латентного периода
двигательной реакции составляет около
120 – 220 мсек., а при утомлении, недостатке
кислорода и пр. она может увеличиваться
до 300 – 500 и более миллисекунд. При более
сложных движениях, например выполнении
фехтовальщиком укола в мишень с выпадом
в ответ на световой сигнал, этот период
значительно больше – около 300 – 400 мсек.
и более (см. рис. 48, Б).
оли
секунды и даже целые секунды. Поскольку
проведение импульсов по нервным волокнам
от периферических рецепторов в нервные
центры и от нервных центров к исполнительным
органам занимает сравнительно небольшое
время, общее время от момента нанесения
внешнего раздражения до появления
ответной реакции организма (латентный
период рефлекса) определяется центральным
временем проведения. Величина латентного
периода рефлекса служит важным показателем
функционального состояния нервных
центров и широко используется в практике.
Латентный период двигательной реакции
измеряется от момента подачи сигнала
до момента начала движения (или до
появления электрической активности
соответствующей мышцы). При осуществлении
человеком простых движений в ответ на
внешние сигналы, например нажимание на
кнопку при световом сигнале (рис. 48, А),
длительность латентного периода
двигательной реакции составляет около
120 – 220 мсек., а при утомлении, недостатке
кислорода и пр. она может увеличиваться
до 300 – 500 и более миллисекунд. При более
сложных движениях, например выполнении
фехтовальщиком укола в мишень с выпадом
в ответ на световой сигнал, этот период
значительно больше – около 300 – 400 мсек.
и более (см. рис. 48, Б).
Укорочение латентного периода одной и той же двигательной реакции человека в процессе спортивной тренировки может отражать повышение функционального состояния нервных центров (ускорение проведения) и перестройку нервных цепей, участвующих в передаче нервных импульсов (укорочение проводящего пути).
7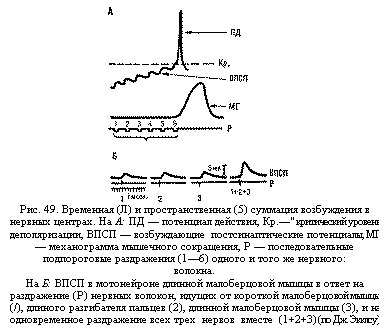 .
Суммация возбуждения. (от позднелат.
summatio — сложение), взаимодействие
синап-тич. процессов (возбуждающих и
тормозных) на мембране нейрона или
мышечной клетки, характеризующееся
усилением эффектов раздражения до
рефлекторной реакции. Явление С. как
характерное свойство нервных центров
впервые описано И.. М. Сеченовым в 1868. На
системном уровне различают С.
пространственную и временную.
Пространственная
С.
обнаруживается в случае одновременного
действия неск. пространственно разделённых
афферентных раздражений, каждое из
к-рых неэффективно для разных рецепторов
одной и той же рецептивной зоны. Временная
С.
состоит во взаимодействии нервных
влияний, приходящих с определ. интервалом
к одним и тем же возбудимым структурам
по одним и тем же нервным каналам. На
клеточном уровне такое разграничение
видов С. не оправдано, поэтому её наз.
пространственно-временной. С. — один
из механизмов осуществления координир.
реакций организма.
.
Суммация возбуждения. (от позднелат.
summatio — сложение), взаимодействие
синап-тич. процессов (возбуждающих и
тормозных) на мембране нейрона или
мышечной клетки, характеризующееся
усилением эффектов раздражения до
рефлекторной реакции. Явление С. как
характерное свойство нервных центров
впервые описано И.. М. Сеченовым в 1868. На
системном уровне различают С.
пространственную и временную.
Пространственная
С.
обнаруживается в случае одновременного
действия неск. пространственно разделённых
афферентных раздражений, каждое из
к-рых неэффективно для разных рецепторов
одной и той же рецептивной зоны. Временная
С.
состоит во взаимодействии нервных
влияний, приходящих с определ. интервалом
к одним и тем же возбудимым структурам
по одним и тем же нервным каналам. На
клеточном уровне такое разграничение
видов С. не оправдано, поэтому её наз.
пространственно-временной. С. — один
из механизмов осуществления координир.
реакций организма.
8. Тонус длительное стойкое возбуждение нервных центров и мышечной ткани, не сопровож дающееся утомлением. Тонус нервных центров называется такое состояние тех или иных отделов головного и спинного мозга, при котором они непрерывно посылают импульсы по соответствующим эфферентным нервам, длительно поддерживая определённое функциональное состояние органов и тканей. Наибольшее значение для организма имеет Тонус центров блуждающего нерва и симпатической нервной системы, регулирующих деятельность сердца, Тонус сосудодвигательных центров и др. Под мышечным Тонус понимают длительное напряжение или сокращение мышц, обеспечивающее поддержание определённой позы и положения тела в пространстве (Тонус скелетных мышц), давления в полости пищеварительных органов, мочевого пузыря, матки, а также кровяного давления (Тонус гладких мышц). Различают контрактильный и пластический Тонус При контрактильном Тонус в мышцах, особенно в скелетных, развивается значит, напряжение, при котором усиливается электрическая активность (потенциалы действия мышц) и отмечается некоторое повышение обмена веществ. В мышцах беспозвоночных и некоторых низших позвоночных контрактильный Тонус складывается по типу тетануса, состоящего из очень медленных и редких волн сокращения, накладывающихся друг на друга. В скелетных мышцах позвоночных контрактильный Тонус поддерживается путём попеременных сокращений отдельных мышечных волокон, входящих в состав мышцы. При пластическом Тонус развиваемое мышцей напряжение невелико, но может поддерживаться длительно без утомления и без значительного повышения обмена веществ. При этом мышца приобретает свойство пластичности, т. е. может значительно растягиваться без одновременного увеличения её упругих свойств. Пластический Тонус основан на длительном слитном возбуждении в мышце, впервые изученном и описанном Н. Е. Введенским. Сопротивление растягивающему усилию во время пластического Тонус осуществляется не столько за счёт возрастающей упругости мышцы, сколько за счёт так называемых вязких сопротивлений, т. е. внутреннего трения. После удаления растягивающей силы мышца не укорачивается до исходной величины, а остаётся более или менее удлинённой; для возвращения её к исходной длине необходимо наличие возбуждающего фактора. Тонус скелетных мышц связан с состоянием мотонейронов спинного мозга, которое зависит от импульсов, поступающих как из вышележащих центров, так и от рецепторов мышц и сухожилий (см. Проприорецепторы). Увеличение афферентной импульсации от мышечных веретён повышает активность мотонейронов спинного мозга и является одной из причин усиления рефлекторного Тонус скелетной мускулатуры. Уровень возбудимости некоторых видов мотонейронов, участвующих в поддержании мышечного Тонус, регулируется ретикулярной формацией ствола мозга. В целостном организме мышечный Тонус поддерживается при участии различных отделов центральной нервной системы. Тоническое напряжение мышц измеряют тонометрами. Нарушение нормальной деятельности нервных центров может сопровождаться как усилением Тонус (гипертония), так и ослаблением его (гипотония и атония). Так, при перерезке мозгового ствола на границе между межуточным и средним мозгом у млекопитающих происходит значительное усиление пластического Тонус; при перерезке на уровне среднего мозга возникает резкое усиление контрактильного Тонус, так называемая децеребрационная ригидность. Аналогичные явления, а также атония возникают и при некоторых заболеваниях центральной нервной системы.
9. Трансформация ритма и силы импульсов. Лат. transformatio - преобразование, превращение - одно из свойств проведения возбуждения в центре, заключающееся в способности нейрона изменять ритм приходящих импульсов. Особенно четко проявляется трансформация ритма возбуждения при раздражении афферентного волокна одиночными импульсами. На такой импульс нейрон отвечает серией импульсов. Это обусловлено возникновением длительного возбуждающего постсинаптического потенциала, на фоне которого развивается несколько ликов (спайков). Другой причиной возникновения множественного разряда импульсов являются следовые колебания мембранного потенциала. Когда его величина достаточно велика, следовые колебания могут привести к достижению критического уровня деполяризации мембраны и обусловливают появление вторичных спайков. В нервных центрах может происходить и трансформация силы импульсов: слабые импульсы усиливаются, а сильные ослабевают.
Окклюзия - нарушение проходимости некоторых полых образований в организме (кровеносных и лимфатических сосудов, подпаутинных пространств и цистерн), обусловленное стойким закрытием их просвета на какомлибо участке.
Острые окклюзии артерий - острое нарушение кровообращения дистальнее места окклюзии артерии эмболом или тромбом. Состояние считают неотложным. Проксимальнее и дистальнее участка окклюзии нарушается нормальный ток кров, что приводит к дополнительному тромбообразованию. Состояние считают обратимым в течение 4-6 часов от его начала (в англоязычной литературе этот временной промежуток называют "золотым периодом"). По истечении этоговремени глубокая ишемия ведёт к необратимым некротическим изменениям.
Окклюзия вены сетчатки - нарушение кровообращения в центральной вене сетчатки или её ветвях. Преобладающий возраст старше 40-50 лет.
Окклюзия центральной артерии сетчатки - острое нарушение кровообращения в центральной артерии сетчатки или её ветвях. Преобладающий возраст 40-70 лет. Преобладающий пол - мужской.