

Общие свойства рефлексов.
Несмотря на все многообразие, рефлексы, отражая, прежде всего, принцип работы нервной системы, обладают рядом общих свойств. Однако, следует помнить, что в силу упрощенного строения рефлекторной дуги, моносинаптические рефлексы, не проявляют многих свойств, напротив, условные рефлексы, имеющие сложную, комбинированную рефлекторную дугу, лежащую в основе сложных форм поведения и научения, дополнительно обладают рядом собственных свойств, что будет рассмотрено в соответствующих разделах.
Поэтому, рассматриваемые нами здесь свойства, в полном объеме будут проявляться только у полисинаптических безусловных рефлексов. Кроме того, лежащие в основе этих свойств физиологические механизмы, объясняющие их проявление и обеспечивающие их реализацию в центральной нервной системе, будут объяснены в материале следующего занятия.
Проведение возбуждения по рефлекторной дуге осуществляется только в одну сторону (от рецептора к центру, от центра к эффектору). Это обеспечивается свойствами химических синапсов.
На развитие рефлекса затрачивается некоторое время, называемое латентным периодом рефлекса или временем рефлекса.
Время рефлекса — это длительность интервала от начала стимуляции до окончания ответной реакции. Оно включает:
время трансформации энергии раздражения в распространяющийся импульс, что осуществляется с помощью рецептора;
время проведения возбуждения в афферентном пути, в центральной части рефлекторной дуги и в эфферентном пути;
время активации эффектора и его ответной реакции.
Время рефлекса зависит, прежде всего, от числа нейронов в рефлекторной дуге, а точнее числа синапсов, так как именно синаптическая задержка является причиной большого времени рефлекса. Поэтому, в моносинаптических рефлексах, типа коленного, у человека время рефлекса практически постоянно и измеряется долями секунды. В этих рефлексах оно не превышает 1,2 мс и в основном создается именно синаптической задержкой.
В полисинаптических рефлексах время рефлекса больше, при умеренных раздражениях оно может измеряться секундами. Более того, время полисинаптических рефлексов непостоянно и может колебаться в достаточно широких пределах. Такое замедление ответа определяется не столько числом синапсов в рефлекторной дуге, сколько тем, что при умеренной силе раздражения активируется небольшое число рецепторов и их импульсы, поначалу, вызывают лишь подпороговые ВПСП в соответствующих вставочных нейронах спинного мозга. Требуется некоторое время на то, чтобы в результате суммации последовательных ВПСП здесь мог быть достигнут порог генерации ПД. И чем меньше сила или время действия раздражителя, тем больше времени на это понадобится.
Кроме того, в интактном организме, на состояние центрального звена рефлекторной дуги полисинаптического рефлекса, могут оказывать влияние нейроны других отделов ЦНС, оказывая тормозящее или активирующее воздействие и, тем самым, влияя на скорость распространения возбуждения.
В центральной части рефлекторной дуги ритм входных (афферентных) сигналов, как правило, трансформируется в иной ритм выходных сигналов. Другими словами, частота импульсов приходящих по афферентному нейрону от рецептора и частота импульсов выходящих по эфферентному нейрону к эффектору, как правило, не одинаково. Это связано с феноменами последовательной суммации ВПСП и с другими причинами, например с явлениями возвратного торможения мотонейронов через клетки Реншоу у позвоночных.
Сила рефлекторною ответа, в конечном счете, определяется числом активированных эффекторных нейронов и частотой их ипульсации.
Сила рефлекса, как правило, растет с усилением раздражения рецепторов. Пределом этого роста является вовлечение в ответ всех мотонейронов в максимальном ритме, хотя предел, обычно, не достигается в связи с подключением других рецепторов (болевых) и возникновением тормозных влияний.
Полисинаптическим рефлексам свойственно последействие т. е. продолжение ответа некоторое время после того, как внешнее раздражение прекращено.
В основе последействия полисинаптического рефлекса лежат во-первых большая длительность ВПСП вставочных нейронов, порождающих не одиночный ПД, а их серию, которая растягивается на десятки и сотни миллисекунд, и во-вторых, дисперсия во времени сигналов, проходящих по параллельным цепочкам нейронов разной сложности и с разными скоростями срабатывания.
В случае очень длительных последействий у рефлексов высшего порядка, например у ориентировочных рефлексов на звуки с задержкой животного в позе прислушивания на секунды и минуты, можно в качестве причины такого последействия предполагать и циркуляцию импульсов в замкнутых нейронных цепях. У моносинаптических рефлексов последействия, как правило, нет.
Многие рефлекторные центры обладают свойством постактивационной потенциации, т. е. усиления ответов на одиночные тестирующие сенсорные сигналы в течение некоторого времени после окончания ритмической активности.
Причина этой потенциации, длящейся несколько минут, лежит в синаптическом облегчении, видимо, отражающем накопление остаточного Са2+ в пресинаптических терминалях при ритмической активности.
В центрах рефлексов может наблюдаться феномен габитуации - ослабления реакции, «привыкания» к раздражителю. Это может быть связано с синаптической депрессией.
В заключение необходимо указать, что центральным звеньям рефлекторных дуг, как правило, свойственна меньшая надежность функции, чем нервным проводникам.
Рефлекторные центры более, нежели проводники страдают от действия различных неблагоприятных факторов, в частности гипоксии и многих фармакологических агентов, например алкоголя, никотина, наркотиков. Особенно ранимы в этом отношении центры полисинаптических рефлексов. Это определяется свойствами синаптических аппаратов вставочных нейронов.
Частная физиология
Рефлексы спинного мозга.
Рефлексы продолговатого мозга
Рефлексы среднего мозга
Рефлексы промежуточного мозга
Мозжечок.
Базальные ганглии.
Спинной мозг.
Спинной мозг является наиболее древним и примитивным отделом центральной нервной системы.
Спинной мозг лежит в позвоночном канале и представляет собой тяж длиной 41 - 45 см (у взрослого), несколько сплющенный спереди назад. Вверху он непосредственно переходит в головной мозг, а внизу заканчивается заострением - мозговым конусом - на уровне II поясничного позвонка.
Характерной чертой организации спинного мозга является периодичность его структуры в форме сегментов, имеющих входы в виде задних корешков, клеточную массу нейронов (серое вещество) и выходы в виде задних корешков. Морфологической границы между сегментами спинного мозга не существует, поэтому деление на сегменты является функциональным. Каждый сегмент через свои корешки иннервирует три метамера тела собственный, половину вышележащего и половину нижележащего, а каждый метамер тела получает иннервацию от трех сегментов спинного мозга. Такое устройство гарантирует осуществление функций спинного мозга при возможных его перерывах и других поражениях.
Распределение функций входящих и выходящих волокон спинного мозга подчиняется определенному закону: все чувствительные (афферентные) волокна входят в спинной мозг через его задние корешки, а двигательные и вегетативные (эфферентные) выходят через передние корешки. Этот закон известен как закон Белла-Мажанди.
Выдающийся шотландский анатом и физиолог Белл и французский исследователь Мажанди установили, что при односторонней перерезке передних корешков спинного мозга отмечается паралич конечностей этой же стороны, чувствительность же сохраняется полностью. Перерезка задних корешков приводит к утрате чувствительности, двигательная функция при этом сохраняется. Таким образом, было показано, что афферентные импульсы поступают в спинной мозг через задние корешки (чувствительные), эфферентные импульсы выходят через передние корешки (двигательные). Эта закономерность известна как закон Белла-Мажанди.
В задних корешках волокон гораздо больше, чем в передних (их соотношение у человека примерно 5:1) т. е. при большом разнообразии поступающей информации организм использует незначительное количество исполнительных приборов. По задним корешкам в спинной мозг поступают импульсы от рецепторов скелетных мышц, сухожилий, кожи, сосудов, внутренних органов. Передние корешки содержат волокна к скелетным мышцам и вегетативным ганглиям.
Задние корешки образованы волокнами одного из отростков афферентных нейронов, тела которых расположены вне центральной нервной системы — в межпозвоночных ганглиях, а волокна другого отростка связаны с рецептором. Общее число афферентных волокон у человека достигает примерно 1 млн. Они различаются по диаметру. Наиболее толстые идут от рецепторов мышц и сухожилий, средние по толщине—от тактильных рецепторов кожи, от части мышечных рецепторов и от рецепторов внутренних органов (мочевого пузыря, желудка, кишечника и др.), наиболее тонкие миелинизированные и немиелинизированные волокна—от болевых рецепторов и терморецепторов. Одна часть афферентных волокон заканчивается на нейронах спинного мозга, другая часть направляется к нейронам продолговатого мозга, образуя спинно-бульбарный путь.
Передние корешки состоят из отростков мотонейронов передних рогов спинного мозга и нейронов боковых рогов. Волокна первых направляются к скелетной мускулатуре, а волокна вторых переключаются в вегетативных ганглиях на другие нейроны и иннервируют внутренние органы.
Спинной мозг имеет два утолщения: шейное и поясничное, соответствующие местам выхода из него нервов, идущих к верхней и нижней конечностям. Передней срединной щелью и задней срединной бороздкой спинной мозг делится на две симметричные половины, каждая в свою очередь имеет по две слабовыраженные продольные борозды, из которых выходят передние и задние корешки - спинномозговые нервы. Эти борозды разделяют каждую половину на три продольных тяжа - канатика: передний, боковой и задний. В поясничном отделе корешки идут параллельно концевой нити и образуют пучок, носящий название конского хвоста.
Внутри спинной мозг состоит из серого и белого вещества. Серое вещество заложено внутри и со всех сторон окружено белым. В каждой из половин спинного мозга оно образует два неправильной формы вертикальных тяжа с передними и задними выступами - столбами, соединенных перемычкой - центральным промежуточным веществом, в середине которого заложен центральный канал, проходящий вдоль спинного мозга и содержащий спинномозговую жидкость. В грудном и верхнем поясничном отделах имеются также боковые выступы серого вещества. Таким образом, в спинном мозге различают три парных столба серого вещества: передний, боковой и задний, которые на поперечном разрезе спинного мозга носят название переднего, бокового и заднего рогов.
П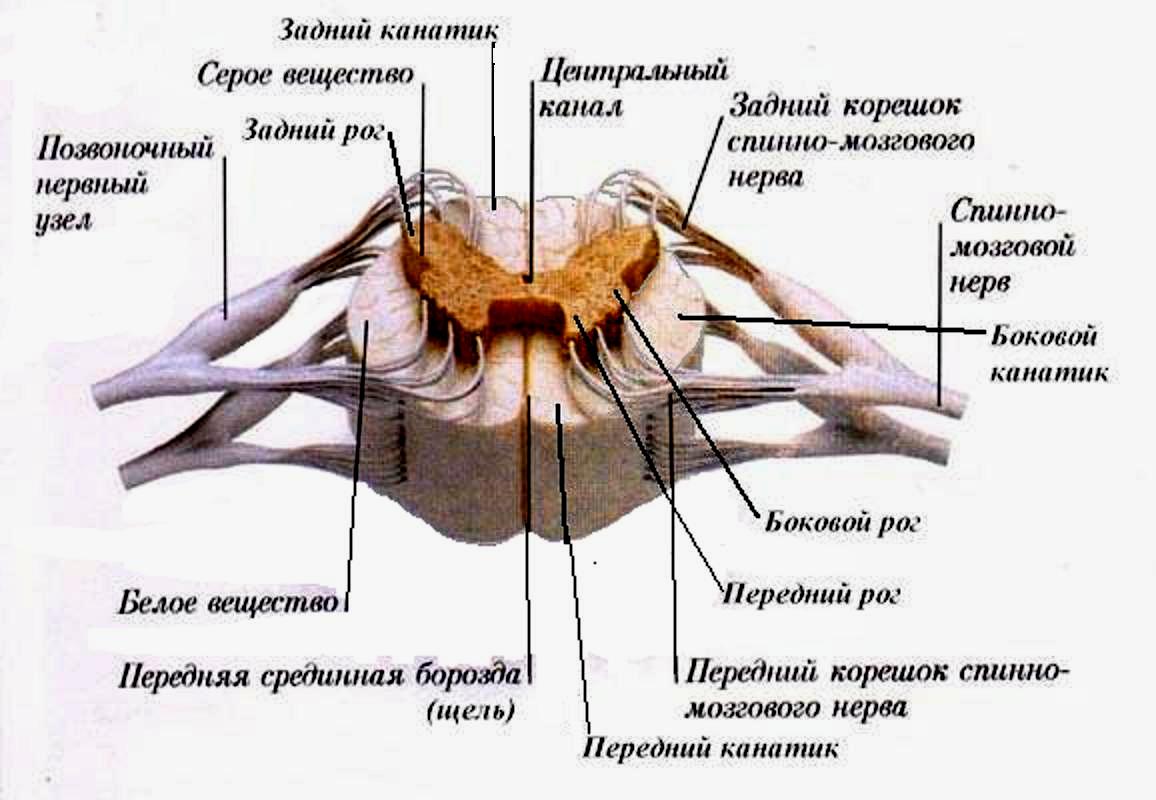 ередний
рог имеет округлую или четырехугольную
форму и содержит клетки, дающие
начало передним (двигательным) корешкам
спинного мозга.
ередний
рог имеет округлую или четырехугольную
форму и содержит клетки, дающие
начало передним (двигательным) корешкам
спинного мозга.
Задний рог уже и длиннее и включает клетки, к которым подходят чувствительные волокна задних корешков.
Боковой рог образует небольшой треугольной формы выступ, состоящий из клеток, относящихся к вегетативной части нервной системы.
Белое вещество спинного мозга составляет передний, боковой и задний канатики и образовано преимущественно продольно идущими нервными волокнами, объединенными в пучки - проводящие пути.
Среди них выделяют три основных вида:
волокна, соединяющие участки спинного мозга на различных уровнях;
двигательные (нисходящие) волокна, идущие из головного мозга в спинной на соединение с клетками, дающими начало передним двигательным корешкам;
чувствительные (восходящие) волокна, которые частично являются продолжением волокон задних корешков, частично отростками клеток спинного мозга и восходят кверху к головному мозгу.
От спинного мозга, образуясь из передних и задних корешков, отходит 31 пара смешанных спинномозговых нервов: 8 пар шейных, 12 пар грудных, 5 пар поясничных, 5 пар крестцовых и 1 пара копчиковых. Участок спинного мозга, соответствующий отхождению пары спинномозговых нерввов, называют сегментом спинного мозга. В спинном мозге выделяют 31 сегмент.
Функции и центры спинного мозга. Спинной мозг выполняет две функции: рефлекторную и проводниковую.
Важную роль в развитии представлений о рефлекторной деятельности спинного мозга сыграли открытия и обобщения английского физиолога, лауреата Нобелевской премии Чарлза Шерринггона (1859 - 1952).
Объем функций, осуществляемых спинным мозгом, чрезвычайно велик. В шейном отделе спинного мозга находятся центр диафрагмального нерва и центр сужения зрачка, в шейном и грудном отделах - центры мышц верхних конечностей, мышц груди, спины и живота, в поясничном отделе - центры мышц нижних конечностей, в крестцовом отделе - центры мочеиспускания, дефекации и половой деятельности, в боковых рогах грудного и поясничного отделов спинного мозга - центры потоотделения и спинальные сосудодвигательные центры.
В естественных условиях эти рефлексы всегда испытывают влияние высших отделов головного мозга. Степень проявления рефлексов зависит от того, сохраняются ли связи структур спинного мозга со структурами головного мозга. После децеребрации (удаления головного мозга) или спинализации (отделения спинного мозга от головного посредством перерезки) исчезают многие сложные формы активности, создаваемые спинным мозгом. При этом определенное значение принадлежит уровню организации подопытного животного. Например, спинальная лягушка, может сидеть и вырываться, когда ее схватывают, собака же сама не может ни стоять, ни ходить. Причина этого в разобщении спинного мозга и вышележащих структур нарушает. При этом, в частности, исчезают периодические разряды дыхательных мышц, обеспечивающие дыхательные движения, пропадают тонические разряды симпатических нейронов, поддерживающих сосудистый тонус и соответственно артериальное давление
В зависимости от числа нейронов, участвующих в проведении возбуждения, рефлекторные дуги спинного мозга делятся на моносинаптические и полисинаптические. За исключением моносинаптических рефлексов растяжения рефлекторные дуги всех остальных спиномозговых рефлексов являются полисинаптическими.
К числу рефлексов спинного мозга относятся защитные рефлексы, рефлексы на растяжение, мышц-антагонистов, висцеромоторные, вегетативные рефлексы. Эта классификация весьма условна, она указывает только на многообразие рефлекторных ответов. Даже у спинального животного трудно встретить рефлексы, которые относились бы только к одной из названных групп.
Защитные рефлексы запускаются, как правило, с рецепторов кожи, хотя характер ответной реакции сильно зависит от силы и вида раздражителя. Чаще всего конечная реакция выглядит в виде усиления сокращения мышц сгибателей.
Рефлексы растяжения проявляются укорочением мышцы в ответ на ее растяжение.
Основными рецепторами в этом случае служат нервно-мышечные веретена, афферентным звеном чувствительные волокна соматических нервов и дорсальных корешков спинного мозга. Эти рефлекторные дуги чаще всего замыкаются в спинном мозге. Начало и конец рефлекторной дуги связаны с мышцей. Рефлексы наиболее выражены в мышцах-разгибателях. Для того. чтобы организм мог противостоять силе земного притяжения, эти мышцы должны находиться в состоянии тонического напряжения. фиологическое значение этих рефлексов состоит в том, что они участвуют в сохранении статики и положения тела, регулируя степень сокращения мышцы в соответствии с падающими на нее раздражениями.
Рефлексы мышц-антагонистов лежат в основе локомоторных актов и характеризуются тем, что при возбуждении мотонейронов сгибателей одновременно происходит торможение мотонейронов мыщц-разгибателей. При этом в конечности другой стороны наблюдаются обратные явления. В целом это создает правильное чередование противоположных по функциональному значению мышечных сокращений. Механизм, обусловливающий такое чередование активности различных двигательных ядер, например при ходьбе, локализуется в интернейронном аппарате спинного мозга. Вместе с тем для его активации необходимо поступление тонического нисходящего сигнала из двигательных центров головного мозга.
Висцеромоторные рефлексы возникают при возбуждении афферентных волокон внутренних органов и характеризуются появлением двигательных реакций мышц грудной и брюшной стенки, мышц-разгибателей спины. Возникновение этих рефлексов связано с существованием конвергенции висцеральных и соматических аффферентных волокон к одним и тем же интернейронам спинного мозга.
Вегетативные рефлексы заключаются, во-первых, в появлении полисинаптических разрядов в преганглионарных симпатических волокнах в ответ на возбуждение симпатических и соматических чувствительных клеток, во-вторых, в возникновении рефлекторных реакций парасимпатических нейронов в ответ на раздражение чувствительных путей.
Наряду с выполнением собственных рефлекторных реакций нейронные структуры спинного мозга служат аппаратом для реализации большого числа сложных процессов, осуществляемых различными отделами головного мозга.