

Глава 13
МОДЕЛИРОВАНИЕ ОНТОГЕНЕТИЧЕСКОЙ И ЭКОЛОГИЧЕСКОЙ ИЗМЕНЧИВОСТИ ПРИЗНАКОВ
В процессе онтогенеза организма происходит реализация генетической информации в конкретных условиях среды. Особенности этого процесса:
- последовательность, смена и длительность этапов онтогенеза находятся под генетическим и средовым контролем;
- прохождение генотипом определенного этапа развития детерминирует возможные варианты следующих этапов;
- на каждом этапе онтогенеза реализуется только часть генетической информации, работают определенные гены; набор генов, детерминирующих развитие признака, может изменяться в процессе онтогенеза и в различных средовых условиях;
- наследуются не сами признаки, а нормы реакции признаков на изменение условий окружающей среды.
В связи с этим представляют большой интерес количественные методы, описывающие рост растений, отдельных органов в изменяющихся условиях среды. В этой главе рассмотрены математические модели, описывающие рост растений (логистическая функция и др.), модели и биометрико-генетические методы количественной оценки конкуренции растений, а также эколого-генетические модели количественных признаков.
13.1. Биометрический анализ роста растений
Рост любого органа растений описывается в виде S-образной (сигмоидальной) кривой Сакса, которая отражает неравномерность данного процесса в онтогенезе при постоянных внешних условиях и носит общебиологический характер. Среди множества количественных моделей роста растений наибольшее распространение получила логистическая функция
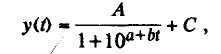 (63)
(63)
где у(t) — величина признака, например, размер (масса) растения или органа; t — время, прошедшее с начала роста; С — начальный размер (масса); А — окончательный прирост к С размера (массы) растения или органа к моменту завершения роста; а, b — константы, определяющие изгиб и наклон логистической кривой.
Логистическая кривая носит симметричный характер, т.е. скорость роста на начальном этапе увеличивается так же быстро, как и падает на заключительном этапе развития растения или органа. Однако не всегда характер роста может быть описан этой кривой. Если рост на начальном этапе замедлен, то кривая приобретает экспоненциальный вид; если же кривая асимметрична в результате замедленного роста на заключительном этапе, то можно воспользоваться т.н. кривой Б. Гомпертца.
И.П.Савиновым с сотрудниками предложена обобщенная формула роста
![]() (64)
(64)
где k — конкретное вещественное число (-<k<+); е — основание натурального логарифма; b<0.
Меняя значение коэффициента k, можно получить любой тип кривой (рис. 38). При k=-1 функция принимает логистический вид, а при k=1 — экспоненциальный и т.д.
 (РИСУНОК
36)
(РИСУНОК
36)
Пример. Воспользуемся данными Г.Н.Зайцева по динамике накопления сухого вещества в плодах томата (точки на рис. 39), для аппроксимации которых он использовал логистическую модель (63).
(РИСУНОК 39)
Через t обозначены пятидневки вегетационного периода, у — содержание сухого вещества в плодах (г. на одно растение). Тогда А=80,3г., С≈0.
Методом наименьших квадратов подобирают константы: а=2,4096, b=-0,2949.
Окончательно теоретическая кривая роста выражается формулой
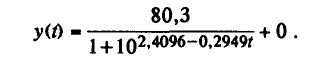
На рис. 39 можно заметить, что эмпирическая и теоретическая кривые довольно близки. Полученную кривую роста сухого вещества в плодах томата можно использовать для прогноза урожая культуры по сухому веществу.
Следует отметить, что тот или иной тип кривой роста, определяемый коэффициентом k в формуле (64), изгиб и наклон кривой, зависящие от констант а и b отражают реализацию генетической программы развития растений в онтогенезе, т.е. являются следствием дифференциальной активности генов. Хотя генетика онтогенеза и в особенности ее количественные аспекты до настоящего времени представляют слабо изученную область биологии, можно проводить селекцию на желательный тип и константы кривой роста, а также на постоянство этих параметров в различных средовых условиях. Возможен и подбор условий среды, в которых кривые роста растений (органов) приближаются к желательному типу.
Тип и параметры кривой не только определяют характер роста растения или органа в онтогенезе при возделывании культур, но и совпадение критических его фаз со сменой лимитирующих факторов среды и в целом конечное значение хозяйственно ценного признака.