

Глава 2
ФАКТОРЫ ИЗМЕНЕНИЯ СТРУКТУРЫ ПОПУЛЯЦИИ
![]()
2.1. Различные схемы скрещивания
Популяционно-генетический анализ не ограничивается панмиксией. Другие схемы скрещивания, встречающиеся в природе и используемые в экспериментах, существенно меняют динамику структуры популяции.
Так при инбридинге скрещиваются особи, родство между которыми в среднем более тесное, чем между особями, случайно взятыми из той же большой популяции. Например, два растения считаются родственниками, если они имеют одного (или более) общего родителя или предка. Сила проявления инбридинга зависит от степени родства скрещивающихся особей.
Существенное значение имеет систематическое родство партнеров скрещивания. При чисто случайном скрещивании в большой популяции родители тоже могут оказаться родственными особями. Но это происходит случайно и не может считаться инбридингом. Популяционно-генетическое последствие инбридинга это прежде всего гомозиготизация, т. е. увеличение долей гомозигот. Крайняя ситуация — инбридинг, вызванный самоопылением всех растений популяции. Особый случай — инбридинг, возникающий в малой по размерам, хотя и скрещивающейся случайно популяции. Здесь вероятность скрещивания родственников увеличивается именно вследствие малого числа особей, участвующих в размножении.
Количественно-генетическое следствие инбридинга и гомозиготизации — инбредная депрессия, проявляющаяся у растений-перекрестников и животных. Поэтому стараются избегать инбридинга используя, в частности, системы самонесовместимости.
Анализ генотипической структуры при инбридинге. При регулярном самоопылении в большой популяции с исходным генотипическим составом f2 (например, 0,25), f1 (0,5), f0 (0,25) доли гетерозигот и гомозигот в поколениях самоопыления изменяются в соответствии с формулами, представленными в таблице 3.
3. Влияние самоопыления на динамику частот генотипов для одного локуса с двумя аллелями
-
Поколение самоопы-
ления
Частота генотипов
Сумма
АА
Аа
аа
I0
f2
(0,25)
f1
(0,5)
f0
(0,25)
1
I1
f2+1/4f1
(0,375)
1/2f1
(0,25)
f0+1/4f1
(0,375)
1
I2
f2+1/4f1+1/8f1
(0,4375)
1/4f1
(0,125)
f0+1/4f1+1/8f1
(0,4375)
1
.
.
.
.
.
.
.
.
.
It
f2+1/2
 1/2)t
f1
1/2)t
f1(1/2)tf1
f0+1/2 1/2)t f1
1
.
.
.
.
.
.
.
.
.
I∞
f2+1/2f1=p (0,5)
0
f0+1/2f1=q (0,5)
1
Итак, при самоопылении частота гетерозигот уменьшается в 2 раза за поколение и приближается к 0, а частоты гомозигот — к частотам р и q аллелей, входящих в них. Сами частоты аллелей не меняются в ряду поколений самоопыления.
Рассмотрим влияние инбридинга на популяцию с исходной генотипической структурой p2, 2рq, q2. При любой схеме скрещивания с инбридингом долю гетерозигот в поколении t популяции удобно обозначать через 2pq×(1-Ft). Ft называется коэффициентом инбридинга. Он оценивает увеличение вероятности появления в локусе двух одинаковых аллелей по причине родственности скрещиваний. Если Ft=0, то доля гетерозигот в поколениях неизменна и составляет 2pq (равновесие по закону Харди-Вайнберга). Если же в поколении t коэффициент инбридинга достиг единицы, то гетерозигот в популяции больше не осталось (f1=0).
При инбридинге снижение доли гетерозигот на 2pqFt вызывает увеличение доли каждой из двух гомозигот на pqFt. Поэтому структура популяции может быть представлена следующим образом:
Генотип АA Аа аа
Частота p2+pqFt 2pq(1-Ft) q2+pqFt.
В научной литературе используется также величина λ, свидетельствующая об уменьшении доли гетерозигот в двух соседних поколениях t и t+1.
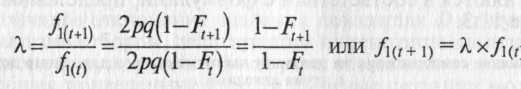
В зависимости от степени инбридинга значение λ, может меняться от 1/2 до 1. При самоопылении λ=1/2 и доля гетерозигот максимально (в 2 раза) уменьшается в каждом последующем поколении. Из последней формулы несложно вывести связь Ft с Ft+1 в следующем поколении самоопыления: Ft+l=1/2(l+Ft). Отсюда, если, например, исходный коэффициент F0=0, легко получить значения Ft в последующих поколениях:
t |
0 |
1 |
2 |
3 |
4 |
5 |
… |
|
|
|
|
|
|
|
… |
Ft |
0 |
1/2 |
3/4 |
7/8 |
15/16 |
31/32 |
… |
Генотип АA Aа аа
Частота p2+pqF3=0,37 2рq(1-F3)=0,06 q2+pqF3=0,57
У животных и двудомных растений сильнейшими формами инбридинга служит постоянное скрещивание между сибсами, а также между родителем (♀ или ♂) и его потомством (♂ или ♀). При таких схемах скрещивания λ после некоторых колебаний в поколениях стабилизируется на уровне λ=0,809. Можно доказать, что после стабилизации λ доля гетерозигот в популяции сокращается приблизительно в 3 раза медленнее, чем в случае регулярного самоопыления. То есть для того же уровня снижения доли f1 нужно в 3 раза меньше поколений самоопыления, чем поколений скрещивания сибсов.
Самоопыление, естественное для пшеницы, ячменя, овса, риса, гороха, табака и т.д., применяется также у перекрестников для получения инбредных линий с целью создания гибридных сортов или ускоренного вытеснения нежелательного рецессивного аллеля.
Неполное самоопыление. Большинство культур не являются исключительно перекрестниками или только самоопылителями. Если в каждом поколении из K-й доли яйцеклеток развиваются семена от перекрестного опыления, а из доли 1-K — семена от самоопыления, то через несколько поколений достигается равновесие генотипического состава популяции. В зависимости от K равновесная доля гетерозигот находится ближе к 2pq или к 0. Ноль будет только при 100%-ном самоопылении (K=0), 2pq при строго перекрестном опылении (K=1). Можно показать, что равновесный состав популяции при определенном K будет:
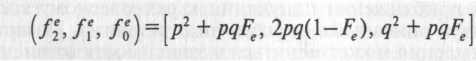
где
равновесный коэффициент инбридинга
определяют по формуле Fe=![]() .
.
Например, при 30%-ном перекрестном опылении (K=0,3) Fe=0,54. Тогда при р=q=0,5 оценка равновесной генотипической структуры, к которой от исходного неравновесия в ряду поколений приблизится популяция, окажется следующей (f2e, f1e, f0e)=(0,385; 0,23; 0,385).
График зависимости Fe от K немного отклоняется вниз от прямой линии (рис. 4).