

Потенциал действия (пд), методы регистрации, фазы, механизмы их происхождении, физиологическое значение
При пороговой силе раздражения в клетке возникает ПД, существенно отличающийся по форме от ЛО (рис.4,Б,1 V).
Он обладает следующими свойствами:
1) подчиняется закону «всё или ничего», т.е. при достижении КУД клетка отвечает максимальным ответом;
2) способен распространяться на большие расстояниям
3) При его возникновении возбудимость клетки снижается;
4) является ауторегенеративным (самоподдерживающимся) процессом.
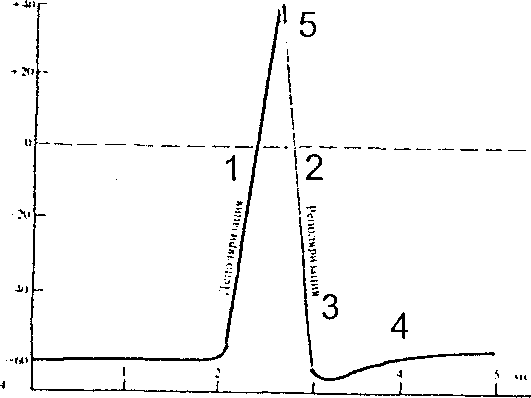

Рис.5. А. фаза потенциала действия: 1- деполяризация, 2- реполяризация, 3- следовая реполяризация, 4- следовая гиперполяризация, 5 - овершут, Б - ионные механизмы развития потенциала действия.
Методика регистрации ПД показана на рис.4,А: при этом один микроэлектрод является раздражающим (1), а второй (2) - отводящим ПД.
ПД имеет достаточно сложную структуру; в нём различают следующие
фазы (рис.5,А):
фаза деполяризации (ЛО не показан);
фаза реполяризации;
следовой деполяризационный потенциал;
следовой гиперполяризационный потенциал;
фаза овершута.
Происхождение этих фаз:
во время фазы деполяризации открываются Na+ -каналы и ионы натрий лавинообразно входят в клетку (рис.5,Б)
во время фазы реполяризации Na+ - каналы закрываются, по открываются К+ - каналы и он выходит из клетки наружу;
во время фазы следовой реполяризации выход К+ несколько замедляется;
во время следовой гиперполяризации часть К+ - каналов открыта и при достижении величины МП калий еще продолжает входить в клетку;
фаза овершута (перевёртывания) – в эту фазу цитоплазма клетки заряжена положительно из-за наличия в ней большого количества ионов Na+.
ПД больше не получается, чем МП: его амплитуда получается при алгебраическом сложении амплитуд овершута и МП; на рис.6,А амплитуда ПД составляет 100 мВ, длительность 1 мс.
Физиологическая роль ПД: возбуждение клеток и возникновение в них соответствующих процессов, передача возбуждения в ЦНС, к периферическим структурам.
Изменения возбудимости клетки во время развития пд. Периоды рефрактерности, механизмы их происхождения, физиологическое значение
В исходном состоянии, когда мембранный потенциал не изменён (рис.6,1;а) возбудимость клетки называется исходной (рис.6,II;а) и составляет 100 %. При возникновении локального ответа (рис.6,I;б), возбудимость клетки повышена (рис.6,II;б). Это связано с уменьшением КУД. При развитии быстрых компонентов ПД (фазы деполяризации и реполяризации - рис.6,I,в) клетка проходит через стадию абсолютной и относительной рефрактерности (рис.6,II,в).
В фазу абсолютной рефрактерности клетка не отвечает на любые, даже сверхсильные раздражения - возбудимость ткани равна нулю. Время этого состояния соответствует длительности фазы овершута (рис.6,I).
В фазу относительной рефрактерности ткань можно возбудить, но более сильными, чем обычно, раздражениями.
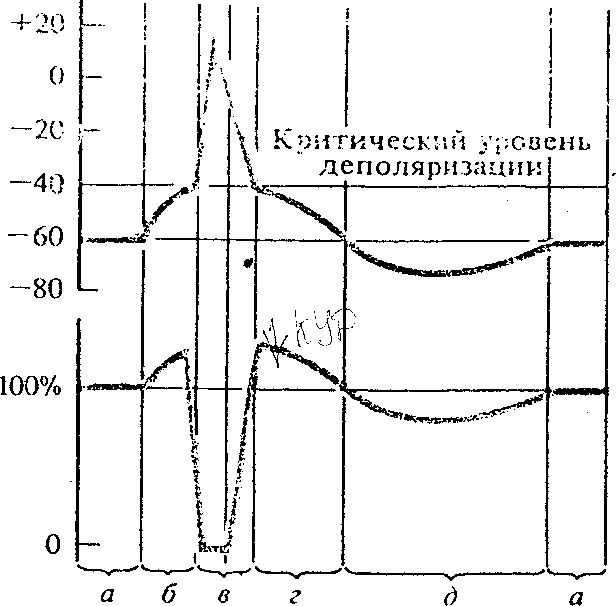
Рис.6. Сопоставление фаз потенциала действия (I) с фазами возбудимости (II). а- исходная возбудимость; б- повышенная возбудимость; в- относительная и абсолютная (О) рефрактерность; г — супернормальная возбудимость; д- субнормальиая возбудимость.
Абсолютная рефрактерность связана с инактивацией - каналов и повышением проводи мости для К+ - ионов. Фазу относительной рефрактерности: первая - связана с постепенной инактивацией Na+- проводимости, вторая - с повышением К+ -проводимости.
В фазу следового деполяризационного потенциала (рис.7, I.г)
возбудимость снова превышает нормальную - т.н. «супернормальная возбудимость» (рис.6,II,г); связана с уменьшением критического уровня деполяризации.
В фазу следовой гиперполяризации (рнс.6,1;д) возбудимость ткани несколько снижена - фаза субнормальной возбудимости (рис.7, II;д). Она снижена из-за повышения КУД.
После восстановления мембранного потенциала ( рис.6,1 ;а) нормализуется и возбудимость (рис.7,11 ;а).
Физиологическое значение изменений возбудимости:
полностью или почти полностью ограждает возбудимую ткань во время возбуждения от посторонних помех (абсолютная и относительная рефрактерность);
повышение возбудимости в фазу ЛО способствует процессам интеграции нейронов в ЦНС;
субнормальная возбудимость в фазу следовой гиперполяризации способствует «отдыху» ткани и восстановлению ионных градиентов клеток.