

5-ші ТАРАУ. АДАПТИВТІ ИММУНИТТІҢ ЖАСУШАЛЫҚ МЕХАНИЗМДЕРІ
Адаптивті иммунитеттің Т-жасушалық звеносы, оның қызметтері. Т-лимфоциттердiң антигентәуелсiз дифференцировкасы. Т-лимфоциттердiң тимустағы оң және терiс сұрыпталуы, олардың маңызы. Реттегіш Т-лимфоциттердің түзілуі. «Аңқау» және эффекторлық Т-лимфоциттер. Т-лимфоциттермен антигендердің танылуы. CD4+Т-хелперлердiң антигентәуелдi дифференцировкасы. Тх1 мен Тх2 функционалды ерекшелiктерi. Тх17, олардың дифференцировкасы мен қызметтері. Реттегіш CD4+Т-хелперлер, олардың субпопуляциясы және қызметтері. CD8+ цитотоксикалық Т-лимфоциттер дифференцировкасы, олардың әсер ету механизмі. Т-зерде жасушалары. Супрессор-жасушаларының адаптивті иммунды жауаптағы маңызы. Маманданған лимфоциттер субпопуляциясы (NK-жасушалары, NKT-жасушалары, γδТ-лимфоциттер) және олардың қызметі.
Адаптивті иммунитеттің жасушалық иммуните жүйесі (Т-жүйесіне) адаптивті иммуниттің бір бөлігі болып табылады. Оған ізашар Т-лимфоциттердің негізгі дамитын тимус (адаптивті иммунитеттің Т-жасушалық звеносының орталық мүшесі), адаптивті иммунитет жүйесінің шеткі мүшелерінде басымырақ түрде Т-лимфоциттер орналасатын аймақтар (Т-тәуелді аймақтар), Т-лимфоциттердің әртүрлі функционалды субпопуляциялары (Т-хелперлер, цитотоксикалық және реттегіш лимфоциттер), сонымен қатар осы жасушалармен өндірілетін цитокиндер (лимфокиндер) кіреді.
Тимуста Т-лимфоциттердің негізгі субпопуляцияларының клондары, олардың оң және теріс сұрыпталу үрдістері, Т-жасушаларының функционалдық әртүрлі негізгі екі субпопуляциясы қалыптасады: CD4+ және CD8+ Т-лимфоциттер. Шеткері мүшелерде аңқау Т-лимфоциттер антигендермен кездеседі және басқа иммунды хабарлы жасушалармен әсерлесу кезінде адаптивті иммунды жауаптың қалыптасуына, реттелуіне және оның жүзеге асуына қатысады.
Т-лимфоциттер рециркуляция үрдісіне белсене қатысады, осы үрдістің арқасында адаптивті иммунды жүйенің қызметтерінің интеграциясы, сонымен қатар иммунды бақылау қызметі жүзеге асады
Адаптавті иммунитеттiң Т-жасушалық звеносыныңнегізгі қызметтерi:
Жасушалық түрдегі адаптивті иммунды жауаппен қамтамасыз ету адаптивті иммунитеттің Т-жүйенiң негiзгi қызметi: Т-лимфоциттер генетикалық бөгде жасушаларға (мутантты және iсiк жасушалары, вируспен зақымдалған жасушалар, бөгде трансплантат жасушасы және т.б.) цитотоксикалық әсер етедi. Одан басқа Т-лимфоциттер кейбір антигендерге қарсы фагоцитарлық түрдегі жауапты қалыптастырады;
Т-лимфоциттер табиғатта кездесетiн көпшiлiк антигендердi тануға және жасушалық, гуморалды иммунды жауаптың қалыптасуында басты қызмет атқарады;
Т-лимфоциттер адаптивті иммунды жауапты реттеуде маңызды қызмет атқарады. Бұл үрдіске, ең алдымен, әртүрлі түрдегі иммунологиялық серпілістерді ынталандыратын және тежейтiн Т-хелперлер мен реттегіш Т-лимфоциттер өздерi өндіретін лимфокиндерiнің көмегiмен қатысады;
Т-лимфоциттер иммунологиялық төзiмдiлiктiң қалыптасуына және оның сақталып тұруына қатысады. Иммунологиялық төзiмдiлiк дегенiмiз организмнiң меншiктi антигендерiне қарсы арнайы иммунологиялық ареактивтілік.
Жетiлген эффекторлы Т-лимфоциттер иммунологиялық зерде, яғни организм антигенмен қайтадан кезедескен жағдайда иммунды жауаптың жылдым қалыптасу қызметiн атқарады.
Т-лимфоциттердiң дифференцировкасы
Т-лимфоциттердiң дифференцировка үрдісінде екi негiзгi кезең бар: антигентәуелсiз және антигентәуелдi дифференцировка.
Т-лимфоциттердiң антигентәуелсiз дифференцировкасы тимуста өтедi, оған iзашар Т-лимфоциттердiң (іТ) тимусқа өтуi, тимоциттердiң пролиферациясы, антигентанушы рецепторлардың түзiлуi, Т-лимфоциттердiң клондарының оң және терiс сұрыпталуы (МНС антигендерi бойынша рестрикцияланған клондардың сақталуы және организмнiң өзiнiң антигендерiмен әсерлесуге қабiлетi бар клондардың жойылуы), бiр-бiрiнен қызметi және беткей CD-маркерлерi бойынша ажыратылатын Т-лимфоциттердiң әртүрлi субпопуляцияларының қалыптасуы жатады.
Антигентәуелдi дифференцировка үрдісі кезінде тимустан шыққан жетiлмеген лимфоциттер сәйкес келетін антигендер клондарымен кездеседі және оларды антигентаныстырушы жасушалармен бiрiге анықтау арқылы бiрқатар керектi стимулдарды алады. Нәтижесiнде Т-лимфоциттер көбейедi және жетiлген эффекторлы Т-лимфоциттерге дифференцияланады. (хелперлер, реттегіш Т-лимфоциттер және цитотоксикалық Т-лимфоциттер). Олар сол антигенмен қайта кездескен жағдайда оны өз бетiмен анықтайды және иммунологиялық жауапты қалыптастырады.
Т-лимфоциттердiң антигентәуелсiз дифференцировкасы
Сүйек кемiгiнде түзiлген іТ-лимфоциттер қан ағысымен тимустың қыртысты аймағына келiп орналасады.
Бұл үрдісте тимустың эпителиалды жасушалары түзетiн хемокиндер маңызды қызмет атқарады (хемокиндер iТ-лимфоциттердi тимусқа тартады). iТ-лимфоциттер тимусқа өту үшiн гематотимикалық тосқауылдан өтуi тиiс. Бұл тосқауылдан өту iТ-лимфоциттердiң мембранды молекулалары (протеогикан CD44, гиалурон қышқылының қалдықтары, β-интегрин және басқа) мен жасушааралық матрикс молекулаларының (фибронектин, тосқауыл жасушаларының мембраналық молекулалары) өзара әсерлесуiне негiзделген.
Тимустың субкапсулярлы аймағында iТ-лимфоциттерден жетiлмеген Т-лимфоциттер түзiледi, олар иммунды хабарлы болып тимустан шығу үшiн дифференцировканың бiрнеше сатысынан өтедi.
Тимустың қыртысты қабатында лимфоциттердiң көбею үрдісі өте интенсивтi жүредi. Адамда орта есеппен тәулiгiне шамамен 5х108 тимоциттер түзiледi, ал тимустан осы уақыт арасында шамамен тек 8х106 жасушалар ғана шығады. Яғни, жаңа түзiлген жасушалардың шамамен 3% тимустан шығады. Бұл құбылыстың биологиялық мәнi жақында ғана белгілі болды. Ол меншiктi гистосәйкестiк антигендерiмен әрекеттесуге қабiлетi бар Т-лимфоциттер клондардың сұрыпталуына негiзделген.
iТ-лимфоциттерден жартылай жетiлген Т-лимфоциттердiң тимус ішіндегі түзiлуi үрдісі фенотиптiк Т-жасушалық маркерлер экспрессиясының өзгеруiмен сипатталады. Олардың негiзiнде беткей CD-антигендер (дифференциялаушы антигендер) жатады.
Әртүрлi CD-антигендер дифференцировканың белгiлi кезеңiне, сондай-ақ функционалды әртүрлi Т-лимфоциттердiң субпопуляцияларына тән.
CD-антигендер – күрделi нәруыздардың, гликопротеидтердiң молекулалары, олар лимфоциттердiң плазмалық мембранасында орналасқан. Т-хелперлер үшiн фенотиптiк маркер CD4 антиген болса, цитотоксикалық Т-лимфоциттер үшiн – CD8. Аталған екi нәруыз корецептор қызметiн атқарады және антигендi тану үрдісіне қатысады.
Кортикалды лимфоциттердiң ең алғашқы даму сатыларында CD2 нәруызы пайда болады және ол Т-лимфоциттер үшін жалпы маркер болып есептеледі, сонымен қатар, ол қой эритроциттерiне рецептор қызметiн атқарады. Аталған рецепторды анықтау үшiн қой эритроциттерiмен қойылатын спонтанды розетка түзу әдiсi қолданады (Е-РОК). Бұл әдiстiң көмегiмен Т-лимфоциттердiң жалпы санын анықтауға болады.
Басқа жалпы Т-лимфоцитарлы маркеріне CD3 нәруызы жатады, ол Т-лимфоциттердiң антигентанушы рецептор мен антиген детерминантасының әсерлесуi кезiнде Т-лимфоциттердiң цитоплазмасына белгi беруде маңызды қызмет атқарады.
Т-лимфоциттердiң дифференцировка үрдісі кезiнде жаңа CD-антигендердiң пайда болуымен қатар кейбiр ескi антигендер жойылуы мүмкiн. Сондықтан, CD-антигендер кезеңерекшелiк антигендердiң мысалы бола алады. Яғни, CD-маркерлер бойынша Т-лимфоциттердiң жалпы саны мен олардың субпопуляцияларының санын және жасушалардың жетiлу дәрежесiн анықтауға болады.
Антигентанушы рецептор (ТЖР - Т-жасушалық рецептор) Т-лимфоциттер үшін басқа негiзгi маркер болып табылады, ол да тимустың қыртысты қабатында, тимоциттер дифференцировкасының алғашқы кезеңдерінде пайда болады.
Дифференцировка үрдісі кезіндегі Т-лимфоциттердiң беткей маркерлерiнiң фенотиптiк өзгеруi бiрқатар стимулдар әсерiнен жасушада өзгерiстердің болғанын, яғни белгiлi бір гендердiң қайта құрылуы мен ынталануын, жасушадағы синтетикалық үрдістердiң және жасушаның белгiлi бір функционалды қасиеттерге ие болуын көрсетедi.
Т-лимфоциттердiң дифференцировкасын шақыратын стимулдарға, ең алдымен, тимоциттердің тимустың эпителиалды жасушаларымен, макрофагтармен, дендриттi жасушалармен жасушааралық әсерлесуi және тимоциттердің адгезия молекулаларымен әрекеттесуі, сонымен қатар адгезия молекулаларының өздері жатады.
Бұл үрдісте гуморалды әсерлер де маңызды рөл атқарады. Олар: тимус гормондары мен бiрқатар цитокиндер: ИЛ-3, ИЛ-1, ИЛ-7 және т.б.
Сүйек кемiгiнен тимусқа келген iТ-лимфоциттер бетiнде беткей молекулалардың белгiлi жинағы бар лимфобласт болып табылады, атап айтқанда CD44. Бiрақ дифференцировка маркерлерi CD4 пен CD8 жоқ. Олар тимустың қыртысты қабатының жоғарғы бөлiгi – субкапсулярлы аймаққа орналасады.
iТ-лимфоциттер мен субкапсулярлы аймақтың стромасының әсерлесуі Т-жасушалардың алғашқы арнайы маркері – CD2 нәруызының пайда болуына әкеледi. Аталған маркерi бар (CD2+) тимоциттер мен эпителиалды «тәрбиешi» жасушалар (nurse cells) тығыз байланыста болу салдарынан белсендi түрде көбейе түседі және жаңа нәруыздарды экспрессиялай бастайды. Олар: CD3, CD4, CD8 және ТЖР-дың β-тiзбегi. Олардың фенотипi келесiдей жазылады: CD2+CD3+CD4CD8βТЖР. Енді осы жасушалар тимустың қыртысты қабатына тереңiрек кiре бастайды. Бұл үрдісте маңызды қызметтi тимопоэтин және ИЛ-3 пен ИЛ-7 цитокиндері атқарады.
Қыртысты қабатта ТЖР қадағалайтын гендердiң қайта құрылуы мен ынталануы нәтижесiнде ТЖР-дiң екі тізбегі де ( және тiзбектері) пайда болады. Бұған қоса CD4 пен CD8 нәруыздары толығымен экспрессияланады.
Қыртысты қабаттағы тимоциттер цитоплазмалық өсiндiлерi тармақталған кортикалды эпителиалды жасушалармен тiкелей байланыста болады. Аталған эпителиалды жасушаларда I және II класс МНС молекулалары жақсы экспрессияланған.
Осы жасушааралық әсерлесу тимустың қыртысты қабатында өтетін негізгі сұрыпталу үрдістерін шақырады. Кортикалды тимоциттердiң бетiнде ТЖР-дың толық экспрессияланауы ерекшелiгi бойынша әртүрлi CD4+CD8+ТЖР+ Т-лимфоциттер клондарының дамуына әкеледi.
Рецепторлары МНС-дің меншiктi нәруыздарына сәйкес келмейтін клондар (олардың саны басымырақ) апоптоз (жасушалардың жоспар бойынша жойылуы) нәтижесiнде жойылады. Тек МНС-дің меншiктi нәруыздарына сай рецепторлары бар клондар сақталады. Бұл Т-лимфоциттер керектi дифференцировка белгiсiн алғандықтан апоптозға ұшырамайды және ары қарай даму сатысына өтедi. Сонымен, өзiнiң организмiнде жұмыс iстеуге қабiлетi бар (МНС-рестрикция) Т-лимфоциттер клондарының сұрыпталуы өтедi.
Бұл үрдіс оң сұрыпталу деп аталады. Оң сұрыпталудан кейiн кортикалды тимоциттердiң шамамен 5% қалады, олар кортикомедуллярлы аймақ арқылы тимустың милы қабатына жылжиды.
Аталған жасушалардың әрқайсысының немесе I класс МНС нәруыздарымен, немесе II класс МНС нәруыздарымен әсерлесуге қабiлетi бар. Оң сұрыпталу үрдісі кезiнде I класс МНС нәруыздарына сәйкестігі бар тимоциттер CD8 корецепторын сақтап қалады, ал CD4 нәруызының экспрессиясын тоқтатады. Бұл жасушалар цитотоксикалық Т-киллерлердiң фенотипiне ие болады: CD2+3+8+ (CD8+).
II класс МНС нәруыздарына сәйкестігі бар тимоциттер CD4 корецепторын сақтап қалады, ал CD8 нәруызының экспрессиясын тоқтатады. Бұл жасушалар Т-хелперлердiң фенотипiне ие болады CD2+3+4+ (CD4+). Осылайша Т-лимфоциттердiң екi негiзгi функционалды субпопуляциялары қалыптасады.
Тимустың кортикомедуллярлы және милы қабаттарында оң сұрыпталудан сақталып қалған Т-лимфоциттердiң клондары бiрқатар цитокиндер мен тимикалық гормондардың (ең алдымен тимозин әсерiнен), сондай-ақ жасушааралық әсерлесу әсерiнен пiсiп жетiлудiң келесi кезеңiне өтедi және терiс сұрыпталу үрдісiне ұшырайды.
Қалыпты жағдайда организмнің иммундық жүйесi меншiктi (аутологиялық) антигендерге төзiмдi. Соңғы көзқарастар бойынша тимустағы терiс сұрыпталу үрдісі нәтижесiнде меншiктi антигендерге төзiмдiлiк қалыптасады деп саналады.
Эксперименталды мәлiметтерге сүйене отырып көптеген зерттеушiлер келесi тұжырымға келдi: тимустың кортикомедуллярлы және милы қабаттарында түзiлген, бiрақ толық жетiлмеген CD4+ және CD8+ Т-лимфоциттер организмнiң меншiктi антигендерiн (фагоцитоз бен пиноцитоздан кейiнгі тимустың меншiктi жасушаларының ыдыраған өнімдері және тимусқа қан арқылы түскен аутологиялық нәруызды молекулалар) иммуногендi түрде, яғни I және II класс МНС нәруыздарымен комплекс түрінде «ұсынып» тұрған макрофагтармен және дендриттi жасушалармен әсерлеседi. Бiрақ, Т-жасушалық рецептор көмегімен Т-лимфоциттер аталған антигендердi анықтаған кезде тек арнайы белгiнi алады, ал апоптоздан қорғану үшiн, олар екiншi – костимулдаушы белгiнi алуы тиiс. Сонымен қатар, тимоциттердiң бетiнде Bcl-2 немесе Bcl-XL нәруыздарының экспрессиясы болуы керек, олар сәйкес онкогендердiң өнiмдерi болып табылады және жасушаны апоптоздан қорғайды. Жетiлмеген тимоциттердiң бетiнде аталған нәруыздар және костимулдаушы молекулалар болмағандықтан немесе олар өте аз мөлшерде болуы салдарынан меншiктi антигендерімен әсерлескен Т-лимфоциттердiң клондары апоптозға ұшырайды.
Сонымен, жасушааралық әсерлесу (костимулдаушы молекулалармен әрекеттесу арқылы) немесе гуморалды өсушi факторлар түрінде қажетті белгiлердiң болмауы нәтижесiнде Т-лимфоциттер тимуста пiсiп жетiлудiң барлық кезеңдерiнде апоптозға ұшырайды.
Клоналды сұрыпталу кезеңдерінде апоптоз керексiз, оң сұрыпталудан өтпеген (МНС-рестрикция) және аутореактивтi клондардың (терiс сұрыпталу) жойылуында маңызды орын алады. Нәтижесiнде меншiктi антигендерге ерекшелiгi жоғары, Т-жасушалық рецепторы бар Т-лимфоциттер клондарының көпшiлiгi жойылады. Қан айналымына Т-жасушалық рецептордың «өзiнiң» антигендерiне ұқсастығы жоқ Т-лимфоциттердiң клондары шығады және сақталады. Бүгінгі күні клонды делеция (терiс сұрыпталу) табиғи иммунологиялық төзiмдiлiктiң түзiлуiнiң негiзгi механизмi деген пiкiр қалыптасқан.
1990-шы жылрадың аяғында 2000-ші жылдардың басында СD4+T-лимфоциттердің жаңа субпопуляциясы ашылды. Олар тимустағы теріс сұрыпталу нәтижесінде, аутореактивті СD4+T-лимфоциттердің альтернативті клонды делециясы ретінде дифференцацияланады және оларда меншікті антигендерге ТЖР аффинділік дәрежесі жоғары болады. Бұл жасушалар АТЖ бетіндегі антигендермен әсерлескенде, транскрипция факторы FoxP3 (ол Т-лимфоциттердің дифференцировкасы және олармен цитокиндер синтезіне жауапты гендердің транскрипциясын реттейді) деңгейі жоғарылайды, CD25 нәруызының (ИЛ-2 арналған рецептор) экспрессиясы жоғарылайды және олар CD4+25+FoxP3+ фенотипті реттегіш Т-лимфоциттерге (Трет.) дифферцацияланады. Олар шеткері мүшелерде жұмыс атқаратын және дәл сол арнайылықтағы жетілген эффекторлық Т-лимфоциттер қатысты супрессорлық белсенділігі бар (Tx1, CD8+ЦТЛ).
Көпшілік Трет.лимфоциттер аутореактивті жасушалар, олар эффекторлы Т-лимфоциттер қатысатын аутоиммунды үрдістерді тежейды. Осының арқасында Трет.лимфоциттер шеткері төзімділік механизмдерінде маңызды орын алады. Мұны жануарларда жүргізілген тәжірибелердің нәтижелері дәлелдейді. Тәжірибеде CD4+25+FoxP3+ фенотипті Т-лимфоциттер аллергиялық энцефаломиелит, эксперименталды аутоиммунды колит, эксперименталды аутоиммунды диабет сияқты аутоиммунды аурулардың дамуын тежеген.
Бiрақ, организмнiң барлық аутоантигендерi тимусқа түспейтінін ескеруіміз керек, сол себептен олар терiс сұрыпталу үрдісіне ұшырамайды. Атап айтқанда, тимусқа көптеген мүшеерекшелiк антигендер түспейдi, яғни аутореактивтi клондардың белгiлi бір саны тимуста жойылу үрдісіне ұшырамайды, нәтижесiнде олар бiрден шеткерi мүшелерге барады. Қалыпты жағдайда бұл клондардың жұмысы төзiмдiлiктiң шеткерi механизмдерi арқылы тежелеуі мүмкін деп болжанады.
Терiс сұрыпталудан сақталған CD4+ және CD8+ Т-лимфоциттер клондары тимустан шығып шеткерi лимфоидты мүшелердiң тимустәуелдi аймақтарына барады, мұнда сәйкес антигендермен кездесу нәтижесiнде антигентәуелдi дифференцировкаға ұшырайды.
Тимустағы оң және теріс сұрыпталудан сақталған CD4+ және CD8+ Т-лимфоциттер, сондай-ақ CD4+25+FoxP3+ Трет. лимфоциттер клондары әлi функционалды жетiлмеген жасушалар болып табылады. Бұл жасушаларды антигендермен кездеспегендіктен «аңқау» лимфоциттер деп атайды. Кейде жетiлген эффекторлы лимфоциттердiң iзашарлары деп те атайды.
Т-лимфоциттердің антигентәуелсіз дифференцировкасы 28-ші суретте көрсетілген.
2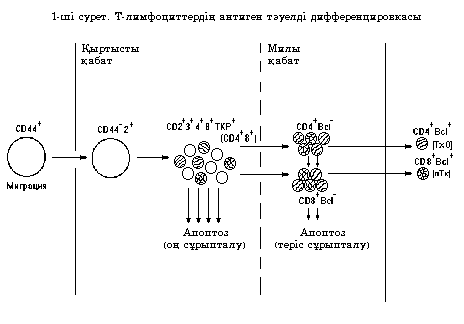
Тимустан шыққан CD4+ Т-лимфоциттер эффекторлы Т-хелперлердің және CD4+25+FoxP3+ Трет. лимфоциттердің, ал CD8+ Т-лимфоциттер эффекторлы цитотоксикалық Т-лимфоциттердің ізашарлары болып саналады (CD8+ ЦТЛ).
Т-лимфоциттердiң антигентәуелдi дифференцировкасы
Тимуста түзiлген Т-лимфоциттер қан ағысымен иммунды жүйесiнiң шеткi мүшелерiнiң Т-тәуелдi аймақтарына келедi. Бұл жасушалардың фенотипi немесе CD4+, немесе CD8+. Тек кейбiр жасушалар тобының (шамамен 2%) фенотипi CD4+CD8+ болуы мүмкін - бұл жетiлмеген жасушалар.
Айналымдағы «аңқау» Т-лимфоциттер бірнеше айдан бірнеше жылға дейін өмір сүруі мүмкін және олар үнемі сүйек кемігінің ізашарларымен алмасып отырады. Сонымен қоса, шеткері мүшелерде жойылған жасушалардың орынын пролиферацияға ұшырай отырып (төменгі деңгейде пролиферацияға ұшырайды), үнемі толтырады. Пролиферация деңгейі лимфопения немесе басқа жағдайлар кезінде жоғарылауы мүмкін. Адамда 60 жас шамасында тимуста жаңа Т-лимфоциттердің түзілуі тоқтатылады (тимустың жасқа байланысты инволяциясы), бірақ, айналымдағы «аңқау» Т-лимфоциттердің саны сақталады. Мұндай механизм ТЖР бойынша арнайылығы әртүрлі, соның ішінде жұқпаларды танитын Т-лимфоциттер клонының репертуарының азаюына әкеледі. Бірақ, бұрын кезедескен антигендерге арнайылығы бар зерде-жасушалардың жинақталуы арқасында, бұл жағдай ішінара өтеледі.
«Аңқау» Т-лимфоциттердің сақталып қалуы кем дегенде екі факторға байланысты. Біріншіден, аффинділігі төмен ТЖР «антигенді пептид-МНС нәруызы» кешенімен байланысуы, екіншіден, организмнің көптеген жасушаларымен үнемі түзілетін ИЛ-7 болуы.
Адаптивті иммунитеттің шеткерi мүшелерінде «аңқау» Т-лимфоциттер антигендермен кездеседі және олардың антигентәуелдi дифференцировкасы басталады. Т-лимфоциттердiң антигентәуелдi дифференцировкасы 4 негізгі кезеңнен тұрады:
Антигенді өңдеу мен таныстыруды жүзеге асыратын АТЖ әсерлесе отырып, функционалды жетiлмеген Т-лимфоциттердің (аңқау) антигендерді тануы;
Олардың жетiлген эффекторлы жасушаға дейiн пролиферация мен дифференцировка түрiнде антигенге қарсы серпілісі;
Иммунды жауаптың эффекторлы кезеңіне – антигендi бейтараптау мен жоюға қатысуы;
Т-зерде жасушаларының түзiлуi.
Антиген организмге қандай жолмен түссе де, ол лимфа арқылы жақын орналасқан лимфа түйіндеріне (регинарлы) жылжиды, мұнда оларды МФ, ДЖ немесе В-лимфоциттер жұтып, жасушаішілік өңдеуден (процессинг) өткізеді, сосын І немесе ІІ класс МНС молекулаларымен кешен түрінде АТЖ бетінде таныстырылады (презентация). Тіндерде түзілген немесе орналасқан антигендерді (мысалы, вирусты жұқпа ошағы, қатерлі ісіктер және т.б.) жетілмеген ДЖ туа біткен иммунитет рецепторлары көмегімен (TLR және NOD-рецепторлар) таниды, белсенеді, жетіледі және регинарлы лимфа түйіндеріне миграцияланады. Мұнда МНс нәруыздарымен кешен түзеген антигенді пептидтерді Т-лимфоциттерге таныстырады.
Жетiлмеген Т-лимфоциттердің антигендермен бірінші кездесуі АТЖ бетiнде болады. АТЖ бетінде антигендi детерминанттар (АТЖ-мен антигеннің жасушаішілік өңделуі кезінде түзілген антигенді пептид түріндегі Т-жасушалық эпитоп) иммуногенді кешен түрінде кездеседі.
Жетiлмеген Т-лимфоциттердiң антигентанушы рецепторының (ТЖР) АТЖ бетiндегi иммуногендiк кешенмен байланысуы (ТЖР, CD3, CD4 немесе CD8 молекулаларының қатысуымен) тек бiрiншi - арнайы белгiмен ғана қамтамасыз етедi. Бiрақ жетiлген эффекторлы Т-лимфоциттерге дейін пролиферация мен дифференцировкаға ұшырау үшін екiншi белгi керек. Бұл белгiні ИЛ-1 және басқа цитокиндер қамтамасыз етеді, сондай-ақ АТЖ бетiндегi В7 корецепторының жетiлмеген Т-лимфоциттердiң бетiндегi CD28 нәруызымен әрекеттесуi де қамтамасыз етедi. Екі белгіні де алу клонның пролиферациясы мен дифференцировкасына ықпал етеді. Егер жасуша тек бiрiншi белгiнi алып, ал екiншi белгiнi алмаса, онда апоптозға ұшырайды және жойылады.
Екi белгiнi де алған CD4+ және CD8+ Т-лимфоциттер ИЛ-2 рецепторларды экспрессиялайды және өздерi ИЛ-2 өндіре бастайды. ИЛ-2 жасуша бетiндегi ИЛ-2-рецепторларына әсер ету (аутокриндi) арқылы жетiлмеген Т-лимфоциттердiң көбеюiн және жетiлген эффекторлы жасушаларға дифференцациялануын ынталандырады. Бұл үрдіс 4-5 тәулікке созылады.
Пролиферацияға ұшыраған белсенген Т-лимфоциттер, 5-7 тәулікте өздерінің шыңына жетеді. Бұл кезде CD4+T-хелперлердің саны шамамен 10 000 есе жоғарылайды, ал CD8+T-лимфоциттердің саны одан күштірек жүреді, мысалы вирусты жұқпа кезінде олардың саны 50 000 есе жоғарылайды. Мұндай жылдам көбею лимфа түйіндерінің ұлғаюына, яғни лимфоаденопатиялардың дамуына әкеледі.
Көбейген Т-лимфоциттер патогенді жоятын функционалдық қасиеті бар жетілген эффекторлы лимфоциттерге дифференцацияланады (мысалы, CD4+Т-лимфоциттер белгілі бір субпопуляциясы түзіледі: Тх1, Тх2, Тх17 немесе CD8+ ЦТЛ қалыптасады).
Пролиферация және дифференцировка нәтижесінде түзілген эффекторлы жасушалар патологиялық ошақтарға миграцияланады және оның тікелей бейтараптауы мен жойылуын (CD8+ЦТЛ) немесе фагоцитарлық түрдегі иммунды жауапты (Тх1) шақырады. Эффекторлық CD4+ Т-хелперлердің бір бөлігі лимфа түйіндеріндегі лимфоидты фолликулаларға жылжиды, мұнда олар В-лимфоциттермен әсерлесіп, антиденетүзілуін іске қосады және реттейді (Тх1 мен Тх2 бір бөлігі, оларды фолликулярлы Т-хелперлер деп те атайды).
Шеткері лимфоидты мүшелерден шыққан эффекторлық Т-лимфоциттердің миграция үрдісін және олардың қаннан патологиялық ошақ бар тіндерге өтуін бірқатар адгезия молекулалары (адгезиндер), соның ішінде селектинде, адрессиндер және интегриндер тұқымдастығындағы молекулалар бақылайды. Адгезия молекулаларының әсерлесу нәтижесінде Т-лимфоциттер мүшенің паренхимасына енеді.
Эффекторлы Т-лимфоциттер енді қосымша костимулдаушы белгілер болмаса да, антигенді өз бетімен анықтауға және оған тікелей әсер етуге мүмкіншілігі болады.
Антигенге жауап берген клонның пролиферациясы шаман тыс болмас үшін, ол Т-лимфоциттердің бетіндегі CTLA-4 ингибиторлық рецепторының экспрессиясы арқылы тежеледі. Бұл рецептор Т-жасушаларының цитоплазмасында кездеседі және ТЖР арқылы ативтенгеннен кейін жылдам 24 сағат ішінде жасуша бетінде шығады. CTLA-4 CD28 сияқты В7 байланысады, бірақ оның аффинділігі CD28 қарағанда біршама жоғары. В7/CTLA-4 байланысуы активация белгісін тоқтатады және клонның пролиферациясы тежеледі.
CTLA-4 гені жоқ жануарларда аутоиммунды зақымдалумен жүретін лимфопролиферация дамиды, нәтижесінде жануар өледі.
CTLA-4 экспрессиясынан кешірек Т-жасушаларында тағы бір тежегіш рецепторы – PD-1 (апоптоздың индукциясын шақыратын рецептор) экспрессияланады. Аталған рецептор PD-1L лигандасымен байланысқанда (әртүрлі тіндердегі көптеген жасушаларда төменгі деңгейде экспрессяиланады) ТЖР-дан және В7/CD28-ден активация белгісінің берілуі тежеледі, нәтижесінде Т-жасушалық жауап тоқтайды.
Адаптивті иммунды жауаптың соңғы кезеңінде антигенге жауап берген клонның саны төмендейді, тек антигенарнайы Т-зерде жасушалардың белгілі саны қалады.
Бұл жасушалардың рецепторлық аппараты жақсы дамыған, оларды цитокиндер мен адгезиндерге де рецепторлар экспрессияланған. Сондықтан Т-зерде жасушалары антиген организмге қайта түскен жағдайда оған тікелей жылдам жауап береді. Көпшілік Т-зерде жасушалары хелперлік субпопуляцияға жатады, яғни CD4+ фенотипі бар және ІІ класс МНС нәруыздарымен рестрикцияланған.
Зерде жасушаларының өмiр сүру ұзақтығы бiрнеше айдан бiрнеше жылдарға дейiн созылады. Олардың сақталып қалуын ИЛ-7 қамтамасыз етеді. CD8+ ЦТЛ сақталып қалу үшін ИЛ-7 мен ИЛ-15 қажет. CD8+ Т-лимфоциттер CD4+ Т-лимфоциттерге қарағанда ұзақ өмір сүреді. Мысалы, қызылшаға қарсы вакцинациядан кейін түзілген CD8+ Т-зерде лимфоциттері 30 жылдан артық өмір сүреді. Белгілі бір антигендерге зерде жасушаларының өмір сүру ұзақтығы генетикалық детерминацияланған.
Т-зерде жасушалары организм жұқпалы агенттермен қайталама кездескен жағдайда қабылдамаушылықты, яғни иммунитетті қамтамасыз етедi.
Т-зерде жасушаларының қалыптасуы көптеген вирусты жұқпаларға қарсы вакцинация механизмінің негізін құрайды.
Т-зерде жасушаларының екі түрі ажыратылады: орталық және эффекторлық. Орталық Т-зерде жасушалары антиген организмге қайталап түскенде оны таниды, сосын жаңа эффекторлы Т-лимфоциттер түзу үшін пролиферацияға ұшырайды. Сондықтан антигенге қарсы екіншілік иммунды жауап 3-4 күннен ерте дамымайды.
Эффекторлық Т-зерде жасушалары антигенмен қайталап кездескеннен кейін бірден белсенеді, сондықтан антигенге жауапты онымен қайта кездескеннен кейін бірден жүзеге асырады.
Т-лимфоциттермен антигендердiң танылуы
Егер В-лимфоциттер қандай да бір нәруыздармен байланыспаған бос антигендi детерминантты анықтауға қабiлетi бар болса, Т-лимфоциттер пептидті Т-жасушалық эпитоптарды тек I және II класс МНС нәруыздарымен кешен үрiнде ғана анықтай алады.
Мысалы, CD8+ ЦТЛ бөгде эпитопты тек I класс МНС нәруыздарымен кешен түрiнде ғана таниды. Бұл шектелудi МНС-рестрикция деп атайды. Сонымен CD8+ ЦТЛ I класс МНС нәруыздары бойынша рестрикцияланған.
Т-хелперлер антигендердi II класс МНС нәруыздарымен кешен түрiнде ғана анықтайды, яғни олар II класс МНС нәруыздары бойынш рестрикцияланған. 29-шы суретте Т-лимфоциттердің негізгі екі түрімен антигендердің танылуы берілген. Т-лимфоциттердің антигентанушы кешені (Т-жасушалық рецептор, Т-хелперлерде CD4 нәруызы, CD8+ ЦТЛ CD8 нәруызы және CD3 нәруызды кешені) мен МНС антигендері бойынша рестрикцияланған антигентаныстырушы жасушалар арасындағы қарым-қатынас көрсетілген.
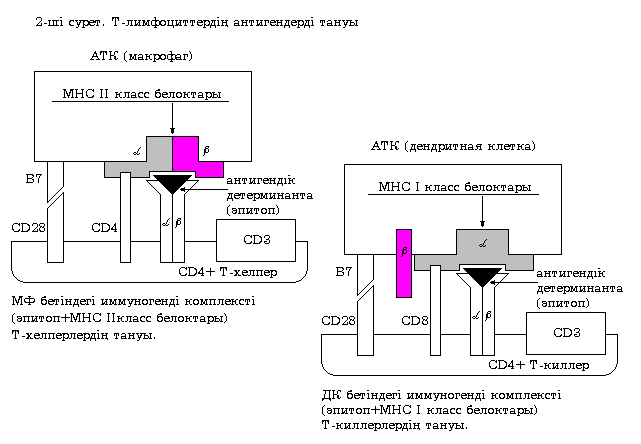

29-шы сурет. Т-лимфоциттермен антигендердің танылуы
Антигентанушы кешен
ТЖР молекулалары бiр-бiрiмен коваленттi байланысқан екi полипептидтi тiзбектен ( және ) тұратын гетеродимер. Тiзбектiң әрқайсысы вариабелдi V-доменi мен тұрақты С-доменiнен тұрады. Вариабелдi домендер антигендi детерминанттармен тiкелей байланысады. Олардың құрамында иммуноглобулиндердiң идиотиптiк детерминанталарына ұқсас идиотиптiк детерминанттар болады.
Т-жасушалық рецептор молекуласының құрылымы иммуноглобулиндер тұқымдастығының құрылымына ұқсас келедi. ТЖР вариабелдi және тұрақты домендерi иммуноглобулиндердiң сәйкес домендерiне ұқсас. Т-жасушалық рецептордың В-лимфоциттердiң иммуноглобулиндi рецепторлардан айырмашылығы – ол бiр валенттi, ал иммуноглобулиндердiң рецепторлары - екi валенттi.
Бүгiнгi күнi Т-жасушалық рецептордың екi түрi белгiлi: ТЖРI және ТЖРII. ТЖРI - және -тiзбектерiнен тұрады және ол эмбрионалды даму кезеңiнде бiрiншi пайда болады, бiрақ ересек организмде аталған рецепторлары бар жасушалар саны 5% аспайды. Бұл субпопуляцияның рөлi әлi толығымен анықталмаған. Т-лимфоциттердiң 95% - және -тiзбектерiнен тұратын ТЖРII экспрессиялайды.
Т-жасушалық рецептордың V-домендерiнiң вариабелдiлiгiн (сәйкес гендердiң мүмкiн болатын барлық комбинациясын ескере отырып) есептеген кезде, Т-жасушалық рецептордың арнайылық варианттары (идиотиптер) шамамен 3х108 құрайтынан көрсеттi.
Вариабелдi домендердiң - және -тiзбектерi антигендi детерминанттармен (эпитоптармен) тiкелей байланысады. Ол антигентаныстырушы жасушамен өңделген антигеннiң антигендi детерминанты мен ТЖР арнайы байланысуын қамтамасыз етедi. Т-жасушалық рецептордың - және тiзбектерiнiң әртүрлi аймақтары АТЖ бетiндегi МНС нәруыздарының белгiлi бiр аймақтарымен және өңделген антигендердiң эпитоптарымен әсерлеседi. Тұрақты домендер МНС нәруыздарымен комплементарлы байланысуын қамтамасыз етедi.
Т-жасушалық рецептор гетеродимерi Т-лимфоциттердiң мембранасында CD3 кешенiнiң полипептидтi тiзбегiнiң цитоплазмалық аймақтарымен ковалентсiз байланысқан. CD3 кешенi антигендi тiкелей тануға қатынаспайды, бiрақ ол ТЖР молекуласынан цитоплазмаға белгiнiң өтуiн жүзеге асырады. Нәтижесiнде жасуша iшiлiк үрдiстер белсену салдарынан Т-лимфоциттердiң эффекторлы қызметiнiң жүзеге асуына мүмкiндiк туады.
Т-хелперлер мен CD8+ ЦТЛ мембранасында орналасқан CD4 және CD8 молекулалары антигендi тану үрдiсi кезiнде АТЖ бетiндегi МНС нәруыздарының сәйкес аймақтарымен (антигендi жасушаiшiлiк өңдеу үрдiсi кезiнде эпитоп пен МНС нәруыздары бiрiгiп иммуногендi түрде болады) әсерлеседi. Бұл әсерлесу антигентаныстырушы кешеннiң жұмысындағы ең маңызды кезеңнiң бiрi болып табылады. Сондықтан CD4 және CD8 молекулаларын Т-жасушалық рецептордың корецепторлары деп атайды.
Антигендердi тану үрдiсi АТЖ (ICAM-1, LFA-3) мен Т-лимфоциттер (LFA-1, LFA-2) бетiндегi арнайы адгезия молекулаларының әрекеттесуi арқылы иницияциаланады және жеңiлденедi.
Т-хелпер мен Т-киллердiң антигендердi тану үрдiсiне толығырақ тоқталсақ.
Антигентаныстырушы жасуша бетiндегi иммуногендi кешендi ТЖР көмегiмен анықтаған кезде Т-хелперлер алған белгi арнайы деп атайды. Бiрақ, Т-хелперлердiң толық белсенуi үшiн бұл белгiнi қабылдау жеткiлiксiз. Сондықтан тағы бiр белгi - костимулдаушы белгi қажет. Егер жасуша аталған белгiнi алмаса, онда апоптозға ұшырайды және жойылады. Т-хелперлер екiншi белгiнi өздерiнiң бетiнде орналасқан CD28 (CD40L немесе CTLA-4) молекуласы мен АТЖ бетiне экспрессияланған костимулятор молекулалары – нәруызды молекулалармен әрекеттескенде алады. Макрофагтар мен дендриттi жасушаларда аталған молекулаларға В7 молекуласы, ал В-лимфоциттерде В40 молекуласы жатады.
Макрофагтарда В7 молекуласының экспрессиясы индуцибелдi. Мысалы, оның экспрессиясын TLR және NOD-рецепторлары арқылы полисахаридтер мен липополисахаридтердiң бактериалды қабырғасы шақыра алады. Бұл қосылыстар В7 мен В40 молекуласының В-лимфоциттерде экспрессиясын шақыруы мүмкiн. В7 молекуласының экспрессиясын шақыратын басқа факторлар әлi белгiсiз. Сонымен қатар, CD4+Т-хелперлердiң костимуляциясына гуморалды факторлар да қатысады. Олар: ИЛ-1, ИЛ-12, ИЛ-4, ИЛ-6, ИЛ-23 және басқа цитокиндер.
Туа біткен иммуниет рецепторлары арқылы В7 костимуляторының микробты компоненттерi әсерiнен шақырылуы иммунды жүйенiң организмнiң меншiктi антигендерiн бактериалды антигендерден ажыратуға мүмкiндiк бередi деген болжам бар. Кейбiр нәруыздарға иммундық жауапты олардың құрамына өлтiрiлген микроорганизмдер мен бактериалды қабырғалардың компонентерi бар адъюванттарды қосқанда ғана шақыруға болатыны эксперименталды түрде дәлелденген. Бұл жағдай келесiдей түсiндiрiледi: егер құрамында бактериалды компоненттерi жоқ нәруызды антигендер макрофагтармен жұтылып және таныстырылса, антигендi таныған Т-лимфоцит костимулдаушы белгiнiң болмауы салдарынан инерттi (анергия) жағдайға немесе апоптозға ұшырайды. Ал иммунизациялаушы қоспаға бактериалды қабырғаның компоненттерiн (макрофагтарда В7 молеукласының өндiрiлуiн шақыратын) қосу CD4+Т-хелперлердiң жұмысының толық iске қосылуын қамтамасыз етедi, нәтижесiнде нәруыздарға қарсы антиденелердiң өндiрiлуi басталады. Бұл үрдiс адъюванттардың көмегiнсiз мүмкiн емес.
ДЖ В7 костимуляторы мен I және II класс МНС молекулалары айқын, әрi тұрақты (индуцибелдi емес) экспрессияланған, бірақ В7экспрессиясы төмен. Дендритті жасушалардың TLR арқылы белсенуі коситмулятордың экспрессиясын жоғарылатады, нәтижесінде Т-лимфоциттер костимулдаушы белгіні қабылдайды. Сондықтан дендриттi жасушалармен таныстырылған антигендер CD4+Т-жасушалардың да, CD8+ ЦТЛ-де пролиферациясы мен дифференцировкасын шақыра алады.
«Аңқау» CD8+ ЦТЛ I класс МНС нәруыздарымен кешен түрiндегi сәйкес эпитопты анықтайды және дендриттi жасушада экспрессияланған В7 молекуласынан костимулдаушы белгiнi алады. Нәтижесiнде белсенедi де бiрқатар цитокиндердi, соның iшiнде ИЛ-2 мен IFN- өндiре бастайды. Аталған цитокиндер Т-киллердiң меншiктi рецепторлары көмегiмен аутокриндi әсер ету арқылы пролиферация мен дифференцировканы ынталандырады, нәтижесiнде жетiлген эффекторлы жасушалар (иммунды киллерлер) түзiледi.
Нәруызды антигендердiң пептидтi фрагменттерi немесе вирустардың пептидтерi дендриттi жасушаларда II класс МНС нәруыздарымен ассоциациялануы мүмкiн. Мұндай жағдайда ДЖ бетiндегi иммуногендi кешендң CD4+Т-хелперлер анықтайды, нәтижесiнде жасушалық және гуморалдық адаптивтік иммунды жауап қалыптасады.
Т-лимфоциттердiң субпопуляциялары және олардың қызметтерi
Т-лимфоциттердің келесідей негізгі субпопуляциялары ажыратылады: CD4+Т-хелперлер, CD4+реттегіш Т-лимфоциттер және CD4+ және CD8+ цитотоксикалық Т-лимфоциттер.
CD4+Т-хелперлер
Т-хелпер субпопуляциясының негiзгi маркерiне CD4 дифференцациялаушы антигенi жатады. Тимустан шыққан аңқау CD4+ Т-лимфоциттер нөлдік хелперлер – Тх0 жасушалар субпопуляциясын құрайды. Бұл жасушаларды бұрынғы әдебиеттерде антигенреактивтi жасушалар немесе Т-индукторлар деп атаған.
Өңделген антигендермен бiрiншiлiк танысатын жасушалар - Тх0 жасушалары. Бұл жасушалардан әртүрлi жағдайларға байланысты жетiлген эффекторлы Тх1 және Тх2 түзiледi. Олар дифференцировканы шақырған антигенге қарсы иммунды жауаптың жүзеге асыруына тiкелей қатысатын жасушалар.
Тх0, Тх1 және Тх2 жасушалары негiзгi фенотиптiк маркерлер жиынтығы бойынша бiр-бiрiнен ажыратылмайды. Олардың барлығының бетiнде CD2, CD3, ТЖР және хелпер субпопуляциясының маркерi CD4 болады (соңғы кезде Тх2 жасушасының Тх1 жасушадан айырмашылығы, Тх2 CD30 антигенiн экспрессиялайды деп есептейдi). Бiрақ әртүрлi хелпер субпопуляциясына жататын лимфоциттер әртүрлi лимфокиндердiң жиынтығын өндiредi, сондықтан олар әртүрлi иммунды жауаптың дамуына қатысады.
Тх0 жасушалары антигенмен кездесу сатысында белсенедi де ИЛ-2 және бiраз мөлшерде ИЛ-4 өндiре бастайды. ИЛ-2 осы клондағы Тх0 жасушаларының көбеюiн аутокриндi ынталандырады. Бiрқатар факторларға байланысты Тх0 жасушаларының дифференцировкасы Тх1 немесе Тх2 бағытында жүредi. Дифференцировка бағытын анықтайтын факторлар толығымен зерттелмеген.
Хелперлердiң дифференцировка бағыты антигеннiң табиғатына (бактериялар, вирустар, бөгде нәруыздар және т.б. және олар өз кезегiнде АТЖ түрiн анықтайды), оның физикалық-химиялық сипаттамасына (дисперстiгi, ерiгiштiгi және т.б.), концентрациясына, организмге түсу жолына, мөлшерiне, сондай-ақ организмнiң осы антигенге жауап беруiнiң генетикалық детерминанттығына байланысты.
Маңызды қызметтi микроайналым атқарады: мүше стромасының жасушаларының беткей молекулалары және осы жасушалармен өндірілетiн гуморалды факторлар. Мысалы, лимфа түйiндерiнiң микроайналымы басымырақ түрде Тх1 түзiлуiне ықпал жасаса, шырышты қабаттардың микроайналымы (iшек, тыныс жолдары) Тх2 дамуына ықпал жасайды.
Сондай-ақ дифференцировканың даму бағытын анықтауда маңызды орынды антигентаныстырушы жасушаның түрi мен қызмет ету ерекшелiгi атқарады. Бiрiншiден, олармен өндiрiлетiн цитокиндер спектрiне байланысты. Егер антигендердi таныстыру қызметiн макрофагтар атқарса, онда олар Тх1 түзiлуiне ықпал етедi (30-шы сурет).
TLR арқылы белсенген макрофаг ИЛ-1 мен ИЛ-12 өндiре бастайды. Кейбiр жұқпалы агенттер, әсiресе жасушаiшiлiк бактериялар мен вирустар, лимфа түйiндерiндегi макрофагтар мен дендриттi жасушалармен ИЛ-12 көп мөлшерде өндiрiлуiн ынталандырады. ИЛ-12 NK-жасушаларының белсенуін шақырады, ал олар өз кезегiнде IFN- өндiре бастайды. ИЛ-12 және IFN- әсерiнен Тх0-жасушалары белсенедi және Тх1 бағытында дифференцацияланады. Сонымен бiрге макрофагтармен өндiрiлетiн ИЛ-12 тек Тх1-жасушаларының түзiлуiн ынталандырып қана қоймай, ол Тх2-жасушаларының түзiлуiн тежейдi. NK-жасушаларымен өндiрiлетiн IFN- да Тх2-жасушаларының түзiлуiн тежейдi.
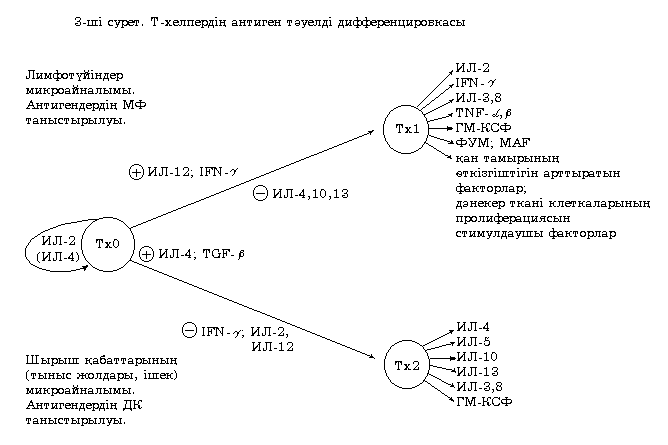

30-шы сурет. Т-хелперлердің антигентәуелді дифференцировкасы
Т-хелпер-2 (Тх2) шырышты қабат микроайналымында Тх0-жасушасына антигендi негiзiнен TLR арқылы белсенген дендриттi жасушалар таныстырғанда түзiледi.
Тх2-дiң түзiлуiн ынталандыратын негiзгi цитокинге ИЛ-4 жатады. Бiраз мөлшерде ИЛ-4 Тх0-жасушалары өндiредi. Сонымен қатар, ИЛ-4 цитокинiн шырышты қабаттардың базофилдерi мен мес жасушалары өндiредi. ИЛ-4 Тх2 түзiлуiне ықпал етумен қатар Тх1 түзiлуiн тежейдi.
Сонымен қатар, Тх2 түзiлуiне шырышты қабаттардың эпителий жасушаларымен өсушi трансформациялаушы фактор (TGF-) әсер етедi. Тх2 белсенуi үшiн ИЛ-1 мен ИЛ-6-ның ынталандырушы әсерi керек.
Тх1 мен Тх2 белгiлi бiр жиынтықтағы цитокиндердi өндiредi, кейбiр цитокиндердi хелперлердiң екi субпопуляциясы да өндiредi. Ал кейбiр цитокиндер тек бiр субпопуляцияға ғана тән болады.
Тх1-жасушалары ИЛ-2 синтездеуiн жалғастырады және IFN-, ИЛ-3 және ИЛ-8 өндiре бастайды. Сондай-ақ гранулоцитарлы-макрофагалды колониестимулдаушы фактор (ГМ-КСФ), iсiкнекроздаушы фактор және (TNF- және TNF-), лейкоциттердiң миграциясын тежейтiн фактор (МТФ), макрофагты қаруландырушы фактор (МАҒ), сонымен қатар дәнекер тiн жасушаларының пролиферациясы мен фибробласттарда коллагеннiң өндiрiлуiн жоғарылататын факторларды өндiре бастайды. Аталған лимфокиндер негiзiнен жасушалық түрдегi адаптивті иммунды жауаптың дамуына ықпал жасайды.
Тх2-жасушалары Тх1-жасушаларынан айырмашылығы, олар ИЛ-2 мен IFN- өндiрмейдi, бiрақ үлкен мөлшерде ИЛ-4 өндiредi. Сонымен қатар, Тх2-жасушалары ИЛ-3, 5, 8, 10, 13 және ГМ-КСФ өндiредi. Қорыта келгенде, ИЛ-3, 8 және ГМ-КСФ Тх1 мен Тх2 үшiн ортақ цитокиндер екендiгiн байқалады.