

6-шы ТАРАУ. МОНОЦИТТЕР/МАКРОФАГТАР ЖӘНЕ ДЕНДРИТТІ ЖАСУШАЛАР, ОЛАРДЫҢ ТУА БІТКЕН ЖӘНЕ АДАПТИВТІ ИММУНДЫ ЖАУАПТАҒЫ МАҢЫЗЫ
Моноциттердің дамуы және тіндік макрофагтар. Резидентті және қабыну макрофагтары, олардың туа біткен иммунды жауапқа қатысу механизмдері. Макрофагтардың антигентаныстырушы қызметі. Дендритті жасушалардың негізгі субпопуляциялары, олардың дамуы және қызметтері. Дендритті жасушалардың антигендерді таныстыру және адаптивті иммунды жауапты іске қосу мен реттеудегі қызметі.
Мононуклеарлы фагоциттер жүйесіне шығу тегі, морфологиясы және қызметтері бойынша біріктірілген шеткері қан моноциттері және әртүрлі орнықтағы макрофагтар кіреді.
Ересек адам организмінде сүйек кемігінде моноцитопоэз үнемі жүреді. Аз жетілген мононуклеарлы фагоциттерге сүйек кемігінің монобласттары жатады. Олар гранулоцитарлық-моноцитарлық ізашар жасушаларынан шығады. Ал олар өз кезегінде көп қабілетті гемопоэтикалық бағана жасушаларынан өрбиді (14-ші сурет).
тін
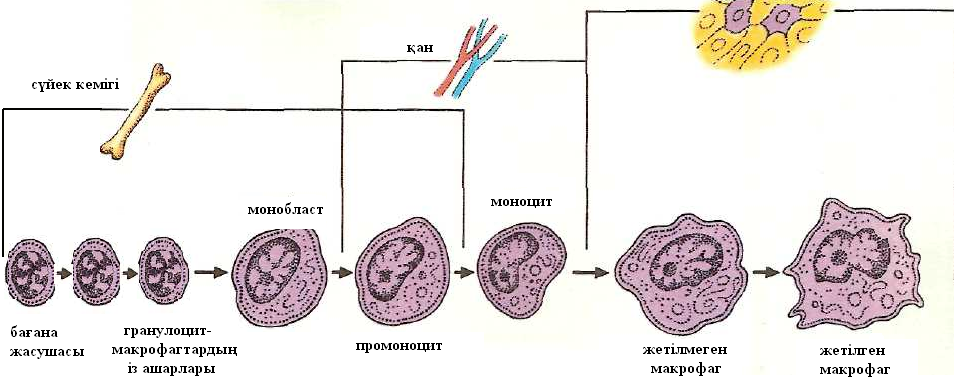
14-ші сурет. Макрофагтар дифференцировкасының негізгі кезеңдері
(В.Г. Галактионов бойынша, Москва, 1986)
Сүйек кемігіндегі моноциттердің өндірілуі бір топ өсу факторларының бақылауында болады. Интерлейкин-3 (ИЛ-3) пен колониестимулдаушы фактор (ГМ-КСФ, М-КСФ) моноциттердің ізашарларының митоздық белсенділігін ынталандырады, ал простагландин Е (PGE) мен интерферондар (IFN-α, IFN-β) осы жасушалардың бөлінуін тежейді.
ИЛ-3 өзінің екінші аты «мульти КСФ» деген атына сәйкес келеді, себебі ол қан жасаудың барлық бұтақтарын ынталандырады, ГМ-КСФ гранулоциттер мен макрофагтардың өндірілуін және макрофагтардың қызметін ынталандырады. М-КСФ мононуклеарлы фагоциттердің арнайы факторы болып есептеледі. Оны сүйек кемігінің стромалды жасушалары, моноциттер, тіндік макрофагтар, Т-хелперлер, фибробласттар, эндометрияның эпителиалды жасушалары және басқа да жасушалар өндіреді. М-КСФ моноциттердің ізашарларының пролиферациясын ынталандырумен қатар олардың микробицидттік және тумороцидтік белсенділігін ынталандырады.
Сүйек кемігінде түзілген моноциттер бір тәуліктен аз мерзімде шеткері қанға шығады. Олардың біраз бөлігі сүйек кемігінде қалып сүйек кемігінің резидентті макрофагтарына айналады. Қабыну кезінде фагоциттеуші жасушалардың қажеттілігі жоғарылауына байланысты, олармен қамтамасыз ету үшін моноциттердің өнімі күрт жоғарылайды. Осыған байланысты промоноциттердің жасушалық цикл уақыты қысқарады, олар көбейе бастайды, ал қанда сәйкесінше моноцитердің саны жоғарылайды. Қабыну ошағындағы макрофагтармен өндірілетін және бөлінетін қабыну алды цитокиндері моноцитопоэзды күшейтетін фактор ретінде болады. Осылайша моноцитопоэдың реттелуі позитивті кері байланыс арқылы жүзеге асады.
Қанда моноциттер қабырғалық және айналымдық деп бөлінеді. Олардың ара-қатынасы ауысып отыруы мүмкін. Айналымдағы моноциттер қан ағымында болғандықтан, қанды зерттеу мақсатында алған жағдайларда, олар зерттеу үлгілеріне түседі, ал қабырғалық моноциттер тамырлардың эндотелиіне берік жабысқан және олар тамырлардан тіндерге қарай трансэндотелиалды миграцияға дайын тұрады. Моноциттердің қаннан тіндерге қарай жылжуы, моноциттер мен эндотелиалды жасушаларда арнайы адгезия молекулаларының болуымен түсіндіріледі. Қан тамырынан шыққан моноцит кері қарай айналымға қайтып келу қабілеті жоқ, ал тіндерге қарай миграция үрдісі үнемі жүреді және ол қабыну кезінде күшейеді, әрі бағытталған сипат алады.
Соңғы жылдары моноциттер популяциясының біркелкі еместігі туралы мәліметтер пайда болды. Мембранды фенотип және функционалды ерекшелігі бойынша моноциттердің екі негізгі субпопуляциясы ажыратылады: CD14++CD16- және CD14+CD16+.
CD14++CD16- субпопуляциясы қан моноциттерінің көпшілігін құрайды. Бұл жасушалардың фагоцитарлық және бактерицидтік белсенділігі жоғары, барлық қабыну алды цитокиндерін түзеді, FcγRI және хемоаттрактанттарға, қабыну хемокиндеріне арналған рецепторлар тығыздығы жоғары. Олардың бетінде β-интегриндер жоғары мөлшерде экспрессияланған. Осындай рецепторлар жиынтығы арқасында моноциттердің бұл субпопуляциясы, бірінші кезекте, қабыну ошақтарының эмирациясы үшін қажет. Бұл аймақтарда олар туа біткен иммунитеттің негізгі қорғаныс серпілістеріне (фагоцитоз, жасушаішілік бактерицидтік және қабынуды дамыту) қатысатын қабыну ошақтарының макрофагтарына айналады.
CD14+CD16+ фенотиптегі моноциттер бетінде жоғары мөлшерде ІІ класс МНС нәруыздары мен костимулдаушы молекулалар экспрессияланған. Олар да фагоцитоз бен қабыну медиаторларын түзуге қатысады, бірақ бұл қызметттер CD14++CD16-моноциттерге қарағанда әлсіздеу. Бірақ CD14+CD16+ моноциттердің антигендерді таныстыру қызметі тиімдірек және олар ИЛ-1 мен IFN-α көп мөлшерде түзеді. Дәл осы жасушалар резидентті тіндік макрофагтардың ізашарлары деген болжам бар.
Тіндік макрофагтардың екі түрі бар: резидентті және қабынулық.
Резидентті макрофагтар CD14+CD16+ моноциттердің қаннан тіндерге қарай қабынулық емес, яғни гомеостатикалық хемокиндердің әсерінен болатын спонтанды миграциясы нәтижесінде үнемі түзіледі. Резидентті МФ үшін негізгі хемокинге фракталкин (СХ3CL1) жатады, оған арналған рецепторлар көп мөлшерде CD14+CD16+ моноциттерде экспрессияланған.
Резидентті МФ гомеостатикалық үрдістерге қатысады: алмасу, реттеу және репаративті. Олар зақымдалған, қартайған және апоптозға ұшыраған жасушаларды утилизациу арқылы қабыну мен некроз ошақтарының және жарақаттардың жазылуына ықпал етеді. Резидентті тіндік МФ комплемент тәуелді лизис және эффекторлық Т-лимфоциттердің цитолитикалық әсері нәтижесінде түзілетін иммунды кешендерді және ыдыраған жасушалық құрылымдар өнімдерін де фагоцитозға ұшыратады. Сонымен қатар, кәсіби антигентаныстырушы жасуша болып табылады.
Резидентті макрофагтарға альвеолярлы, плевралды, перитонеалды макрофагтар, бауырдың Купфер жасушалары, дәнекер тінінің гистиоциттері, остеокласттар, микроглия, бүйректің мезингиалды жасушалары, тимустың макрофагтары және шеткері лимфоидты мүшелердің резидентті МФ жатады. Аталған жасушалардың бір-бірінен морфологиясы, беткей рецепторлары мен маркерлері, түзетін цитокиндер жиынтығы және қызметтері бойынша біршама айырмашылықтары бар.
Тіндік резидентті макрофагтар ұзақ өмір сүретін жасушаларға жатады. Олардың өмір сүру уақыты айлар мен жылдарға созылады. Егер жұқпа немесе қабыну ошағында мобилизациясы болмаса, олар көкбауырға немесе лимфа түйіндеріне барып жойылады. Альвеолярлы макрофагтар өкпенден мұрын жолдары арқылы шығады.
Резидентті макрофагтардың көпшілігі дәнекер тінінде, шырышты қабаттармен ассоциацияланған лимфа түйініндері мен лимфоидты тіндерде, соның ішінде мұрын жолдарының шырыштарында (өкпенің интерстициалды макрофагтары) кездеседі.
Альвеолярлық макрофагтар өкпенің иммундық гомеостазын ұстап тұратын ең маңызды жасушалардың бірі болып есептеледі. Плевралды макрофагтар ерекше субпопуляцияны құрайды, олар қасиеттері бойынша перитонеалды макрофагтарға жақын.
Бауырдағы резидентті макрофагтар – құрамында эндогенді пероксидаза мен біріншілік лизосомды ферменттер бар жұлдызшалы Купфер жасушалары, олар фагоцитоз бен пиноцитоз қызметтерін атқарады. Бауырдаң Купфер жасушалары мононуклеарлық фагоциттер жүйесінің барлық жасушаларының 50% құрайды. Олар тамырлар мен гепатоциттер арасындағы кеңістікте орналасады және қан тамырынан бауырға түскен өнімдердің сүзілуіне қатысады.
Орталық жүйке жүйесінде резидентті тіндік макрофагтардың негізгі қызметтерін (фагоцитоз, цитокиндерді өндіру, цитотоксикалық әсер, антигендерді таныстыру және т.б.) микроглия жасушалары мен астроциттер атқарады.
Тимустағы резидентті макрофагтардың маңызды қызметтерінің бірі – тимустың қыртысты қабататында теріс сұрыпталу нәтижесінде жойылатын тимоциттерді жұту мен олардың утилизациясы.
Шеткері қан моноциттері тек тіндік макрофагтарға ғана емс, сондай-ақ белгілі бір факторлардың қатысуымен дендритті жасушаларға (ДЖ) дифференцациялана алады. Мұндай факторлар қызметін ГМ-КСФ пен ИЛ-4 атқаратыны тәжірибеде дәлелденген. Аталған цитокиндердің әсерінен ДЖ мономорфты популяциясы түзіледі, олар сипаттамасы бойынша шеткері тіндердің жетілмеген ДЖ ұқсайды.
Макрофагтардың жетілуі, дифференцировкасы және белсенуі өсу факторларына (ИЛ-3, ГМ-КСФ, М-КСФ) және белсендіруші цитокиндерге (IFN-γ) тәуелді.
IFN-γ қызметтерінің ішіндегі ең маңыздысының бірі – резидентті және қабыну макрофагтарының эффекторлық қызметін белсендіру. Яғни, макрофагтардың жасушаішілік микробицидтік және цитотоксикалық әсерлерін күшейту, олармен цитокиндердің, супероксидтік және нитроксидтік радикалдардың, проcтогландиндердің өнімін жоғарылату. IFN-γ әсерінен І және ІІ класс МНС-антигендерінің экспрессиясы жоғарылайды, осыған байланысты антигендердің таныстырылу тиімділігі және олардың Т-лимфоциттермен танылуы жоғарылайды.
Қабыну тіндік макрофагтары CD14++CD16- моноциттерінен түзіледі, олардың бетінде қабыну хемокиндеріне арналған көптеген рецепторлар бар, бірінші кезекте, CD14++CD16- моноциттерінің маркері болып табылатын CCR2 бар. Аталған маркер резидентті макрофагтарда кездеспейді, сондықтан CCR2 экспрессиясы бар моноциттер қабыну ошақтарына миграцияланады.
Қабыну макрофагтары туа біткен иммунитеттің негізгі эффекторлық жасушаларының бірі, себебі олар көптеген патогендердің фагоцитозы мен жасушаішілік киллингін жүзеге асырады. Қабыну алды цитокиндердің көпшілігін түзе отырып қорғаныс қабыну серпілісінің дамуын ынталандырады және оған туа біткен иммунитет пен адаптивті иммунитеттің басқа да жасушаларынын қосады.
Макрофагтардың рецепторлар
Макрофагтардың бетінде рецепторлардың үлкен жиынтығы бар. Олардың көмегімен макрофагтар көптеген физиологиялық серпілістерге, соның ішінде туа біткен және адаптивті иммунды жауапқа қатысады.
МФ мембранасында көптеген патогендердің ПАМҚ тануын және жасуша өміріне қауіпті әсерлермен және ситуациялармен ассоциацияланған молекулалық құрылымдарды – ҚАМҚ тануын, бірінші кезекте стрестік нәруыздардың танылуын қамтамасыз ететін туа біткен иммунитеттің паттерн-танушы рецепторлары экспрессияланған.
МН/МФ жетекші ПТР қатарына Толл-тәріздес және NOD-рецепторлары жатады. Бұл жасушалардың бетінде плазмалық жасушалардың мембранасында экспрессияланатын барлық белгілі TLR кездеседі. Олар: TLR1, TLR2, TLR4, TLR5, TLR6 және TLR10. Цитоплазмада жасушаішілік TLR3, TLR7, TLR8, TLR9, сондай-ақ, NOD1 және NOD2-рецепторлары. ПТР репертуарының мұндай кең болуы МФ-мен патогендердің барлық түрлерін (бактериялар, вирустар, саңрауқұлақтар, қарапайымдар, паразиттер) тануға мүмкіндік береді, сонымен қатар патологиялық өзгерген, зақымдалған және апоптоздық жасушаларға тән молекулалық құрылымдарды таниды. II бөлім 2-ші тарау «Туа біткен иммунитет рецепторлары және олармен танылатын молекулалық құрылымдар» қараңыз.
МФ мембраналық CD14 нәруызының «бактериалды ЛПС-ЛПС-байланыстырушы нәруыз» кешенімен байланысуы ЛПС TLR4 байланысуын жеңілдетеді.
Моноциттердің бетінде ПТР жататын аминопептидаза N (CD13) бар, бірақ МФ кездеспейді. CD13 молекуласының кейбір вирус қабығындағы нәруыздарды байланыстыру қабілеті бар.
МН/МФ көптенген фагоцитарлық рецепторларды экспрессиялайды. Олар лектинді рецепторлар (бірінші кезекте маннозалық рецептор, дектин-1 және DS-SING) және патогендер мен басқа да фагоцитоз объектілерін тікелей танитын скавенджер-рецепторлар (II бөлім 2-ші тарау «Туа біткен иммунитет рецепторлары және олармен танылатын молекулалық құрылымдар» қараңыз) бар. Кейбір бактериялардың компоненттері, атап айтсақ, стафилококк, нейссерия, листерия, сонымен қатар өзгерген меншікті жасушалар, модификацияланған төменгі тығыздықтағы липопротеиндер және апоптоз жасушаларының фрагменттері скавенджер-рецепторлар үшін лигандалар болып табылады.
Маннозалық рецептор арқылы МН/МФ көптеген микроорганизмдерді ұстайды, соның ішінде Mycobacteria, Leismania, Legionela, Pseudomonas aeruginosa және т.б. Бұл рецептордың құрылысы бактериялардың жасушалық қабырғасындағы пепдидогликанмен жоғары аффинді байланысуын қамтамасыз етеді. Ең қызығы, макрофагтардың белсенуін шақыратын цитокиндер (IFN-γ, TNF-α) бұл рецептордың түзілуін тежейді және оның экспрессиясын төмендетеді. Ал керісінше, қабынуға қарсы кортикостероидтар оның өндірілуін және экспрессиясын жоғарылатады. D витамині маннозаға арналған рецептордың экспресиясын ынталандырушы екен.
Макрофагтардың мембранасында гликолизденудің соңғы өнімдерін (AGEs) байланыстыруға арналған рецепторалар анықталған. Олар жас ұлғайған сайын тіндерде жинақтала бастайды, ал диабет кезінде жинақталуы жылдамдайды. Бұл гликолиздену өнімдері нәруыздармен айқаса әреткеттесуі нәтижесінде тіндердің зақымдалуын шақырады. AGEs үшін арнайы рецепторлары бар МФ осы өнімдер нәруыздарын ұстайды және оларды деградацияға ұшыратады. Нәтижесінде тіндер деструкциясы дамуының алды алынады.
МН/МФ барлық фагоцитарлы рецепторлары да экспрессияланған. Олардың көмегімен антиденелермен және комплементпен опсонизацияланған патогендердің, сондай-ақ басқа да бөгде бөлшектер мен жасушалардың танылуы жүзеге асады. Оларға бірінші кезекте Ғс-рецепторлар және белсендірілген комплемент фрагменттері үшін рецепторлар (CR1, CR3 және CR4, С1q фрагменті және С3а мен С5а анафилатоксиндеріне арналған рецепторлар) жатады.
Ғс-рецепторлар антиденелермен опсонизацияланған объектілердің танылуы мен фагоцитозын қамтамасыз етеді.
IgG үшін үш түрлі рецепторлар бар: FcγRI, FcγRII және FcγRIII (сәйкесінше CD64, CD32 және CD16). FcγRI – мономерлі IgG үшін аффинділігі жоғарылығымен сипатталатын жалғыз рецептор және барлық макрофагтарда экспрессияланған. Аффинділігі төмен FcγRII рецептор моноциттер мен макрофагтарда экспрессияланған. FcγRIII моноциттер мен макрофагтарда экспрессияланған, IgG үшін аффинділігі төмен және негізінен иммунды кешендерді немесе агрегацияланған IgG байланыстырады. Рецепторлардың барлық үш түрі IgG-мен опсонизацияланған бактериялардың және басқа жасушалардың фагоцитозын шақырады, мембранасында антиген-антидене кешені бар нысаны-жасушаларға қатысты табиғи киллерлер мен фагоциттердің антиденеге тәуелді жасушалық цитоксикалық (АТЖЦ) механизміне қатысады. Нысана-жасуша ретінде вирустермен, қарапайымдармен зақымдалған жасушалар немесе қатерлі трансформацияға ұшыраған организмнің меншікті жасушалары бола алады. Мұндай қорғаныс варианты кейбір паразитарлық және вирусты жұқпалар кезінде ең тиімді болып саналады, себебі макрофагтар өздігімен патогендердің өзін немесе олардың арнайы антиденелермен түзілген кешенін ұстай (жұта алмайды) алмайды. Ғс-рецепторы арқылы белсенген макрофагтар нысана-жасушаның лизисін бірқатар медиаторларды (бірінші кезекте TNF-) бөлу арқасында жүзеге асырады. Кейбір цитокиндер (IFN-γ мен ГМ-КСФ) моноциттре мен макрофагтардың қатсуымен жүретін АТЖЦ тиімділігін жоғарылату қабілеті бар.
Маңызды рецепторлар тобына хемокиндер мен басқа хемоаттрактанттар үшін рецепторлар жатады. Қабыну немесе жұқпа ошағына МН/МФдың хемотаксисін шақыратын С3а, С5а, С5b67 үшін рецепторлардан басқа, бұл жасушалардың бетінде қабыну хемокиндері үшін рецепторлар бар (CXCR1, CCR1, CCR2, CCR3, CCR4, CCR5, CCR8 және т.б.).
Эпителиалды жасушалармен, қан таымрларының эндотелиалды жасушаларымен, сонымен қатр серпіліс ошағындағы резидентті МФ түзілетін қабыну хемокиндері қорғанысқа қатысатын жаңа жасушалардың хемотаксисін ынталандырады. Қабыну ошағына бірініші болып нейтрофилдер келеді, кейінірек моноцитарлы-макрофагалды инфильтарция басталады. Бұл үрдіс аталған жасушалардағы хемокин рецепторалының сәйкес лигандалармен байланысуы салдарынан дамиды.
МН/МФ-дың мембранасында цитокиндер үшін көптеген гликопротеинді рецепторлар экспрессияланған. Цитокиндердің сәйкес лигандалармен байланысуы белсендіруші белгінің жасуша ядросына берілу тізбегінің алғашқы кезеңі болып табылады. МН/МФ-тар үшін ерекше арнайы рецепторлың бірі – ГМ-КСФ (CD115) үшін рецептор. Аталған рецептордың болуы моноциттер мен олардың ізашарларын гранулоцитарлық қатардағы жасушалардан ажыратуға мүмкіндік береді.
МН/МФ үшін ерекше маңызды рецепторлар қатарына IFN-γ үшін рецептор (IFNγRI және IFNγRII) жатады. Себебі олар арқылы осы жасушалардың көптеген қызметтері белсенеді. Сондай-ақ қабыну жауабына қатысатын МН/МФ-дың аутокринді стимуляциясын шақыратын қабыну алды цитокиндері (ИЛ-1, ИЛ-6, TNF-α, ИЛ-12, ИЛ-18, ГМ-КСФ) үшін де рецепторлар бар.