

4. Аеробна система енергозабезпечення рухової активності
Аеробна система енергозабезпечення значно поступається алактатній і лактатній по потужності енергопродукції, швидкості включення в забезпеченні м'язової діяльності, однак багатократно перевершує по ємності і економічності.
Особливістю аеробної системи є те, що утворення АТФ в клітинних органелах – мітохондріях, що знаходяться в м'язовій тканині і примикають до міофібрил, або розкиданих по саркоплазмі, відбувається за участю кисню, який доставляється киснево-транспортною системою, чим зумовлюється висока економічність аеробної системи, а достатньо великі запаси глікогену в м'язовій тканині й печінці, а також практично необмежені запаси ліпідів у адіпозній та м'язовій тканинах – її ємність.
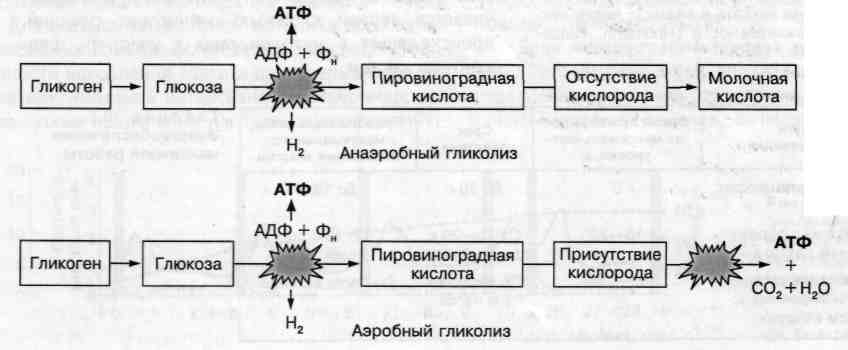
Всі численні хімічні реакції, які виникають в процесі аеробного ресинтезу АТФ, можна розділити на три типи: 1 – аеробний гліколіз; 2 – цикл Кребса, 3 – система транспорту електронів.
Першим етапом реакцій є аеробний гліколіз, в результаті якого здійснюється розщеплення глікогену з утворенням С02 і Н20. Протікання аеробного гліколізу відбувається за тією ж схемою, що і протікання розглянутого вище анаеробного гліколізу. В обох випадках в результаті хімічних реакцій глікоген перетворюється в глюкозу, а глюкоза – в піровиноградну кислоту з ресинтезом АТФ. У цих реакціях кисень не бере участь. Присутність кисню проявляється в подальшому, коли за його участі піровиноградна кислота не перетворюється в молочну кислоту, а потім у лактат, що має місце в процесі анаеробного гліколізу, а направляється в аеробну систему, кінцевими продуктами якого стає вуглекислий газ ( С02), який виведиться з організму легенями, і вода.
З утвореної в результаті розщеплення глікогену піровиноградної кислоти відразу виводиться С02, перетворюючи її з трьовуглецевої сполуки у двовуглецеву, яка поєднуючись з коферментом А, утворює ацетил-КоА, який включається у другий етап реакцій аеробного утворення АТФ – цикл лимонної кислоти або цикл Кребса.
У циклі Кребса протікає серія складних хімічних реакцій, в результаті яких відбувається окислення піровиноградної кислоти – виведення іонів водню (Н+) і електронів (е-), які в результаті потрапляють в систему транспорту кисню і беруть участь в реакціях ресинтезу АТФ на третьому етапі, утворюючи С02, який дифундує в кров і переноситься в легені, з яких і виводиться з організму. У самому циклі Кребса утворюється тільки 2 моля АТФ.
Третій етап реакцій протікає в ланцюзі транспортування електронів (дихального ланцюга). Реакції, що відбуваються за участю коферментів, в загальному вигляді зводяться до наступного. Іони водню і електрони, що виділяються в результаті реакцій, що протікають в циклі Кребса і в меншій мірі в процесі гліколізу, транспортуються до кисню, щоб в результаті утворити воду. Одночасно енергія, яка виділяється в серії сполучених реакцій використовується для ресинтезу АТФ. Весь процес, що відбувається по ланцюгу передачі електронів кисню, називається окислювальним фосфорилюванням.
Проте в цілому глікогенні ресурси м'язів і печінки, можливості глюконеогенезу обмежені і дозволяють забезпечити виділення 1200-2000 ккал енергії (приблизно 5000-8400 кДж), що недостатньо для повноцінного забезпечення тренувальної, а також тривалої змагальної діяльності, характерної для шосейних велосипедних гонок, марафонського бігу, тріатлону та ін. Енергозабезпечення в зазначених видах діяльності багато в чому визначається здатністю організму спортсмена до окислення жирів, запаси і енергоємність яких значно перевершує аналогічні показники глікогену.
Жирові запаси, що звільняються в результаті ліполізу, приблизно в рівній мірі охоплюють два джерела. Перший – ліпозна тканина, з якої в кров надходить близько половини ВЖК(вільних жирних кислот), які доставляються до м'язових кліток для подальшої утилізації. Друге джерело – внутрішньо м'язовий триацилгліцерол, який зберігається в адіпозних клітинах, що знаходяться між волокнами, або у вигляді тріацілгліцеролових утворень безпосередньо у волокнах, і використовуються для розщеплення та подальшої постачання ВЖК в плазму крові.
Хоча в процесі м'язової діяльності ліполіз розгортається поступово, проте вже на 30-40-й хвилинах роботи середньої інтенсивності її енергозабезпечення в рівній мірі здійснюється за рахунок окислення як вуглеводів, так і ліпідів. Подальше продовження роботи, призводить до поступового вичерпання обмежених вуглеводних ресурсів, пов'язано із збільшенням окислення СЖК; наприклад, енергозабезпечення другої половини марафонській дистанції в бігу, або шосейних гонках (більше 100 км) переважно пов'язано з використанням жирів.
Роль білків для ресинтезу АТФ не суттєва. Однак вуглецевий каркас багатьох амінокислот може бути використаний як енергетичне паливо в процесі окислювального метаболізму, що проявляється при тривалих навантаженнях середньої інтенсивності, при яких внесок білкового метаболізму в продукування енергії може досягти 5-6% загальної потреби в енергії. Витрата нескоротливої фракції білків супроводжується звільненням окремих амінокислот, які можуть бути піддані окисленню в скелетних м'язах (Graham et al., 1999). Розпад білків, що відбувається в м'язах при напруженій фізичній роботі, не лише вносить певний внесок у задоволення потреб організму в енергії (Lemon, Mullin, 1980), але і значною мірою зумовлює специфічність м'язової адаптації як реакцію на навантаження різного переважного напрямку (Hickson , Wolinsky, 1989). Згідно з дослідженнми А. Виру (1987) і після закінчення напруженої роботи розпад м'язових білків може продовжуватися, що, однак, слід розглядати не як показник превалюючого розпаду скорочувальних білків, а як відображення рівня їх перетворення, що лежить в основі адаптаційних реакцій, які відбуваються в м'язовій тканині.