

7.6. Цикл кори
Цикл Кори (глюкозо-лактатный цикл) открыла чешская ученая, лауреат Нобелевской премии Тереза Кори. Он представляет собой биохимический транспорт лактата из мышц в печень и дальнейший синтез глюкозы из лактата, катализируемый ферментами глюконеогенеза (рис. 40).
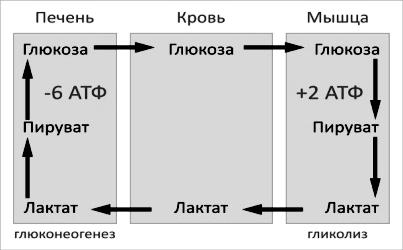
Рис. 40. Схема цикла Кори
При интенсивной мышечной работе и в условиях отсутствия или недостаточного числа митохондрий (например, в эритроцитах) глюкоза подвергается анаэробному гликолизу с образованием лактата. При накоплении лактата в мышцах возникает лактат-ацидоз, раздражаются чувствительные нервные окончания, что вызывает мышечную боль.
Лактат переносится кровью в печень и превращается в пируват, а затем в глюкозу (глюконеогенез), которая с током крови может возвращаться в работающую мышцу.
Направление лактатдегидрогеназной реакции в работающих мышцах и печени обусловлено отношением концентраций восстановленной и окисленной форм НАД+: отношение НАД+/НАДН в сокращающейся мышце больше, чем в печени.
7.7. Спиртовое брожение
Спиртовое брожение осуществляется дрожжеподобными организмами, а также некоторыми плесневыми грибками:
С6Н12О6 2С2Н5ОН + 2СО2
глюкоза этанол
Расхождение гликолиза и спиртового брожения начинается после образования пирувата: при гликолизе пируват при участии лактатдегидрогеназы и кофермента НАДН восстанавливается в лактат, а при спиртовом брожении пируват подвергается декарбоксилированию с образованием ацетальдегида, который восстанавливается в этанол:


При молочнокислом брожении ПВК не декарбоксилируется, а, как и при гликолизе в животных тканях, восстанавливается при участии ЛДГ за счет водорода НАДН.
7.8. Пентозофосфатный путь превращения глюкозы
Пентозофосфатный путь – альтернативный путь окисления глюкозы. Его функции:
- поставляет клеткам кофермент НАДФН, использующийся как донор водорода в реакциях восстановления;
- обеспечивает клетки пентозофосфатами для синтеза нуклеотидов и нуклеиновых кислот.
Пентозофосфатный путь не приводит к синтезу АТФ.
Ферменты пути локализованы в цитозоле.
Пентозофосфатный путь образования пентоз протекает в печени, жировой ткани, молочной железе, коре надпочечников, эритроцитах.
В пентозофосфатном пути превращения глюкозы выделяют окислительный и неокислительный пути образования пентоз.
Окислительный путь включает две реакции дегидрирования. Коферментом дегидрогеназ является НАДФ+, восстанавливающийся в НАДФН. Пентозы образуются при окислительном декарбоксилировании.
Неокислительный путь включает реакции переноса 2- и 3-х углеродных фрагментов с одной молекулы на другую. Этот путь служит для синтеза пентоз. Процесс обратим, и из пентоз могут образовываться гексозы.
1). Дегидрирование глюкозо-6-фосфата при участии глюкозо-6-фос-фатдегидрогеназы и кофермента НАДФ+ с образованием 6-фосфоглюконо--лактона и НАДФН:

2). 6-фосфоглюконо--лактон нестабилен и гидролизуется с образованием 6-фосфоглюконата (фермент – 6-фосфоглюконолактоназа):


3). Дегидрирование и декарбоксилирование 6-фосфоглюконата с образованием рибулозо-5-фосфата (пентоза) и НАДФН при участии декарбоксилирующей 6-фосфоглюконатдегидрогеназы:

4). Под действием эпимеразы из рибулозо-5-фосфата образуется ксилулозо-5-фосфат (пентоза). Под влиянием изомеразы рибулозо-5-фосфат превращается в рибозо-5-фосфат (пентоза). Между формами пентозофосфатов устанавливается равновесие:

На этом этапе пентозофосфатный путь может быть завершен.
При определенных условиях наступает неокислительная стадия пентозофосфатного цикла, протекающая анаэробно: образуются вещества, характерные для гликолиза (фруктозо-6-фосфат, фруктозо-1,6-бисфосфат, фосфотриозы), и вещества, специфические для пентозофосфатного пути (седогептулозо-7-фосфат, пентозо-5-фосфаты, эритрозо-4-фосфат).
5). Транскетолазная реакция взаимодействия ксилулозо-5-фосфата и рибозо-5-фосфата (кофермент – тиаминпирофосфат, переносит гликоальдегидную группу от ксилулозо-5-фосфата к рибозо-5-фосфату):

6). Транскетолазная реакция взаимодействия ксилулозо-5-фосфата и эритрозо-4-фосфата:

7). Трансальдолазная реакция – трансальдолаза катализирует перенос остатка диоксиацетона от седогептулозы-7-фосфата на глицеральдегид-3-фосфат:

Шесть молекул глюкозо-6-фосфата в пентозофосфатном цикле образуют 6 молекул рибулозо-5-фосфата и 6 молекул СО2. Из 6 молекул рибулозо-5-фосфата регенерируется 5 молекул глюкозо-6-фосфата:
6Глюкозо-6-фосфат + 7Н2О + 12НАДФ+
5Глюкозо-6-фосфат + 6СО2 + Pi + 12НАДФН + 12Н+.
Промежуточные продукты цикла (фруктозо-6-фосфат и глицеральдегид-3-фосфат) включаются в гликолиз.