

7.4. Включение фруктозы и галактозы в гликолиз
Фруктоза и галактоза не имеют специфических метаболических путей и через ряд реакций включаются в гликолиз.
Включение фруктозы в метаболизм в печени начинается с реакции фосфорилирования, катализируемой фруктокиназой с образованием фруктозо-1-фосфата:
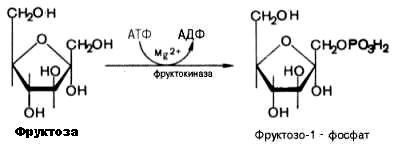
Фруктозо-1-фосфат расщепляется альдолазой на глицеральдегид и диоксиацетонфосфат. Диоксиацетонфосфат включается в пятую реакцию гликолиза.
Глицеральдегид может включаться в гликолиз после его фосфорилирования с участием АТФ. Образовавшийся глицеральдегид-3-фосфат вступает в шестую реакцию гликолиза.
Метаболизм фруктозы в мышечной ткани, почках, жировой ткани начинается с ее фосфорилирования при участии гексокиназы и АТФ. Образуется фруктозо-6-фосфат. Реакция ингибируется глюкозой. Далее фруктозо-6-фосфат превращается во фруктозо-1,6-бисфосфат и включается в четвертую реакцию гликолиза.
Галактоза образуется в кишечнике при гидролизе лактозы. Чтобы превратить галактозу в глюкозу, необходимо провести реакцию эпимеризации. Эта реакция в клетке возможна только с уридиндифосфат-производным галактозы (УДФ-галактоза). Вначале галактоза фосфорилируется, образуется галактозо-1-фосфат. Затем:
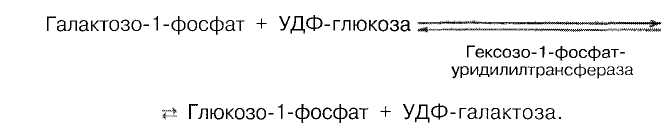
УДФ-галактоза подвергается эпимеризации:


Глюкозо-1-фосфат либо превращается под действием фермента фосфоглюкомутазы в глюкозо-6-фосфат и включается во вторую реакции гликолиза, либо преобразуется в глюкозу при участии фосфатазы:
![]() ,
,
![]() .
.
7.5. Челночные механизмы
Цитозольный НАДH, образующийся в 6 реакции гликолиза, катализируемой глицеральдегидфосфатдегидрогеназой, не может передавать водород в дыхательную цепь, потому что митохондриальная мембрана для него непроницаема. Перенос водорода через мембрану происходит с помощью специальных систем, называемых «челночными». Водород транспортируется через мембрану при участии пар субстратов, связанных соответствующими дегидрогеназами, т.е. с обеих сторон митохондриальной мембраны находится специфическая дегидрогеназа.
Глицеролфосфатная челночная система работает в клетках белых мышц, печени, мозге.
Водород от НАДH в цитозоле передается на диоксиацетонфосфат глицерол-3-фосфатдегидрогеназой (НАД-зависимый фермент) (рис. 38). Образовавшийся глицерол-3-фосфат окисляется ферментом внутренней мембраны митохондрий - глицерол-3-фосфатдегидрогеназой (ФАД-зависимый фермент). Затем протоны и электроны с ФАДH2 переходят на убихинон и далее по дыхательной цепи.
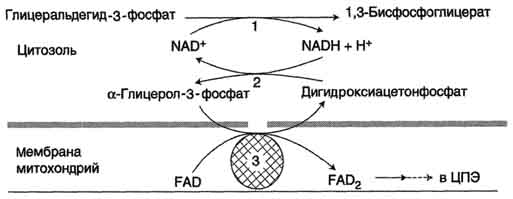
Рис. 38. Глицеролфосфатная челоночная система:
1 - глицеральдегид-3-фосфатдегидрогеназа;
2 - глицерол-3-фосфатдегидрогеназа (цитозольный фермент);
3 - глицерол-3-фосфатдегидрогеназа (митохондриальныи фермент)
Малат-аспартатная челночная система, в которой участвуют малат, цитозольная и митохондриальная малат-дегидрогеназы, является более универсальной, и работает в сердечной мышце, печени, почках.
В цитоплазме НАДH восстанавливает оксалоацетат в малат (реакция 1), который при участии переносчика проходит в митохондрии, где окисляется в оксалоацетат НАД-зависимой малатдегидрогеназой (реакция 2). Восстановленный НАДН отдает водород в митохондриальную ЦПЭ (рис. 39).
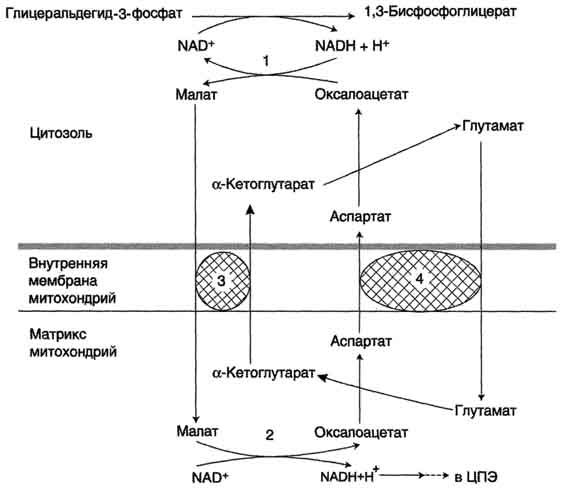
Рис. 39. Малат-аспартатная челоночная система:
1, 2 - окислительно-восстановительные реакции, обеспечивающие транспорт водорода из цитозоля в митохондрии на ЦПЭ;
3, 4 - транслоказы, обеспечивающие транспорт -кетоглутарата, аспартата и глутамата и через мембрану митохондрий
Образованный из малата оксалоацетат выйти из митохондрий в цитозоль не может: мембрана митохондрий для него непроницаема. Поэтому он превращается в аспартат, который транспортируется в цитозоль, где снова превращается в оксалоацетат.
Обе челночные системы отличаются по количеству синтезированной АТФ. В первой системе образуется 2АТФ (водород вводится в ЦПЭ на уровне убихинона). Вторая система энергетически более эффективна – образуется 3АТФ (водород поступает в ЦПЭ с митохондриальным НАД+).