

Прокариотические и эукариотические клетки: строение, отличия Строение эукариотической и прокариотической клеток. Эукариотическая клетка. Строение прокариотической клетки. Сравнение прокариотической и эукариотической клеток. |
||||||||||||||||||||||||||||||||||||
У современных и ископаемых организмов известны два типа клеток: прокариотическая и эукариотическая. Они столь резко различаются по особенностям строения, что это послужило для выделения двух надцарств живого мира - прокариот, т.е. доядерных, и эукариот, т.е. настоящих ядерных организмов. Промежуточные формы между этими крупнейшими таксонами живого пока неизвестны. Основные признаки и отличия прокариотических и эукариотических клеток (таблица):
Основное отличие прокариотических клеток от эукариотических заключается в том, что их ДНК не организована в хромосомы и не окружена ядерной оболочкой. Эукариотические клетки устроены значительно сложнее. Их ДНК, связанная с белком, организована в хромосомы, которые располагаются в особом образовании, по сути самом крупном органоиде клетки - ядре. Кроме того, внеядерное активное содержимое такой клетки разделено на отдельные отсеки с помощью эндоплазматической сети, образованной элементарной мембраной. Эукариотические клетки обычно крупнее прокариотических. Их размеры варьируют от 10 до 100 мкм, тогда как размеры клеток прокариот (различных бактерий, цианобактерий - сине- зеленых водорослей и некоторых других организмов), как правило, не превышают 10 мкм, часто составляя 2-3 мкм. В эукариотической клетке носители генов - хромосомы - находятся в морфологически оформленном ядре, отграниченном от остальной клетки мембраной. В исключительно тонких, прозрачных препаратах живые хромосомы можно видеть с помощью светового микроскопа. Чаще же их изучают на фиксированных и окрашенных препаратах. Хромосомы состоят из ДНК, которая находится в комплексе с белками- гистонами, богатыми аминокислотами аргинином и лизином. Гистоны составляют значительную часть массы хромосом. Эукариотическая клетка имеет разнообразные постоянные внутриклеточные структуры - органоиды (органеллы), отсутствующие в прокариотической клетке. Прокариотические клетки могут делиться на равные части перетяжкой или почковаться, т.е. образовывать дочернюю клетку меньшего размера, чем материнская, но никогда не делятся путем митоза. Клетки эукариотических организмов, напротив, делятся путем митоза (исключая некоторые очень архаичные группы). Хромосомы при этом "расщепляются" продольно (точнее, каждая нить ДНК воспроизводит около себя свое подобие), и их "половинки" - хроматиды (полноценные копии нити ДНК) расходятся группами к противоположным полюсам клетки. Каждая из образующихся затем клеток получает одинаковый набор хромосом. Рибосомы прокариотической клетки резко отличаются от рибосом эукариот по величине. Ряд процессов, свойственных цитоплазме многих эукариотических клеток, - фагоцитоз, пиноцитоз и циклоз (вращательное движение цитоплазмы) - у прокариот не обнаружен. Прокариотической клетке в процессе обмена веществ не требуется аскорбиновая кислота, но эукариотические не могут без нее обходиться. Существенно различаются подвижные формы прокариотических и эукариотических клеток. Прокариоты имеют двигательные приспособления в виде жгутиков или ресничек, состоящих из белка флагеллина. Двигательные приспособления подвижных эукариотических клеток получили название ундулиподиев, закрепляющихся в клетке с помощью особых телец кинетосом. Электронная микроскопия выявила структурное сходство всех ундулиподиев эукариотических организмов и резкие их отличия от жгутиков прокариот 1. Строение эукариотической клетки. Клетки, образующие ткани животных и растений, значительно различаются по форме, размерам и внутреннему строению. Однако все они обнаруживают сходство в главных чертах процессов жизнедеятельности, обмена веществ, в раздражимости, росте, развитии, способности к изменчивости. Клетки всех типов содержат два основных компонента, тесно связанных между собой, — цитоплазму и ядро. Ядро отделено от цитоплазмы пористой мембраной и содержит ядерный сок, хроматин и ядрышко. Полужидкая цитоплазма заполняет всю клетку и пронизана многочисленными канальцами. Снаружи она покрыта цитоплазматической мембраной. В ней имеются специализированные структуры-органоиды, присутствующие в клетке постоянно, и временные образования — включения. Мембранные органоиды: наружная цитоплазматическая мембрана (HЦM), эндоплазматическая сеть (ЭПС), аппарат Гольджи, лизосомы, митохондрии и пластиды. В основе строения всех мембранных органоидов лежит биологическая мембрана. Все мембраны имеют принципиально единый план строения и состоят из двойного слоя фосфолипидов, в который с различных сторон ива разную глубину погружены белковые молекулы. Мембраны органоидов отличаются друг от друга лишь наборами входящих в них белков. Цитоплазматическая мембрана. У всех клеток растений, многоклеточных животных, у простейших и бактерий клеточная мембрана трехслойна: наружный и внутренний слои состоят из молекул белков, средний — из молекул липидов. Она ограничивает цитоплазму от внешней среды, окружает все органоиды клетки и представляет собой универсальную биологическую структуру. В некоторых клетках наружная оболочка образована несколькими мембранами, плотно прилегающими друг к другу. В таких случаях клеточная оболочка становится плотной и упругой и позволяет сохранить форму клетки, как, например, у эвглены и инфузории туфельки. У большинства растительных клеток, помимо мембраны, снаружи имеется еще толстая целлюлозная оболочка — клеточная стенка. Она хорошо различима в обычном световом микроскопе и выполняет опорную функцию за счет жесткого наружного слоя, придающего клеткам четкую форму. На поверхности клеток мембрана образует удлиненные выросты — микроворсинки, складки, впячивания и выпячивания, что во много раз увеличивает всасывающую или выделительную поверхность. С помощью мембранных выростов клетки соединяются друг с другом в тканях и органах многоклеточных организмов, на складках мембран располагаются разнообразные ферменты, участвующие в обмене веществ. Отграничивая клетку от окружающей среды, мембрана регулирует направление диффузии веществ и одновременно осуществляет активный перенос их внутрь клетки (накопление) или наружу (выделение). За счет этих свойств мембраны концентрация ионов калия, кальция, магния, фосфора в цитоплазме выше, а концентрация натрия и хлора ниже, чем в окружающей среде. Через поры наружной мембраны из внешней среды внутрь клетки проникают ионы, вода и мелкие молекулы других веществ. Проникновение в клетку относительно крупных твердых частиц осуществляется путем фагоцитоза (от греч. "фаго” — пожираю, "питое” — клетка)[2]. При этом наружная мембрана в месте контакта с частицей прогибается внутрь клетки, увлекая частицу в глубь цитоплазмы, где она подвергается ферментативному расщеплению. Аналогичным путем в клетку попадают и капли жидких веществ; их поглощение называется пиноцитозом (от греч. "пино” — пью, "цитос” — клетка). Наружная клеточная мембрана выполняет и другие важные биологические функции. Цитоплазма на 85 % состоит из воды, на 10 % — из белков, остальной объем приходится на долю липидов, углеводов, нуклеиновых кислот и минеральных соединений; все эти вещества образуют коллоидный раствор, близкий по консистенции глицерину. Коллоидное вещество клетки в зависимости от ее физиологического состояния и характера воздействия внешней среды имеет свойства и жидкости, и упругого, более плотного тела. Цитоплазма пронизана каналами различной формы и величины, которые получили название эндоплазматической сети. Их стенки представляют собой мембраны, тесно контактирующие со всеми органоидами клетки и составляющие вместе с ними единую функционально-структурную систему для осуществления обмена веществ и энергии и перемещения веществ внутри клетки.
В стенках канальцев располагаются мельчайшие зернышки—гранулы, называемые рибосомами. Такая сеть канальцев называется гранулярной. Рибосомы могут располагаться на поверхности канальцев разрозненно или образуют комплексы из пяти-семи и более рибосом, называемые полисомами. Другие канальцы гранул не содержат, они составляют гладкую эндоплазматическую сеть. На стенках располагаются ферменты, участвующие в синтезе жиров и углеводов. Внутренняя полость канальцев заполнена продуктами жизнедеятельности клетки. Внутриклеточные канальцы, образуя сложную ветвящуюся систему, регулируют перемещение и концентрацию веществ, разделяют различные молекулы органических веществ и этапы их, синтеза. На внутренней и внешней поверхности мембран, богатых ферментами, осуществляется синтез белков, жиров и углеводов, которые либо используются в обмене веществ, либо накапливаются в цитоплазме в качестве включений, либо выводятся наружу. Рибосомы встречаются во всех типах клеток — от бактерий до клеток многоклеточных организмов. Это округлые тельца, состоящие из рибонуклеиновой кислоты (РНК) и белков почти в равном соотношении. В их состав непременно входит магний, присутствие которого поддерживает структуру рибосом. Рибосомы могут быть связаны с мембранами эндоплазматической сети, с наружной клеточной мембраной или свободно лежать в цитоплазме. В них осуществляется синтез белков. Рибосомы кроме цитоплазмы встречаются в ядре клетки. Они образуются в ядрышке и затем поступают в цитоплазму. Комплекс Гольджи в растительных клетках имеет вид отдельных телец, окруженных мембранами. В животных клетках этот органоид представлен цистернами, канальцами и пузырьками. В мембранные трубки комплекса Гольджи из канальцев эндоплазматической сети поступают продукты секреции клетки, где они химически перестраиваются, уплотняются, а затем переходят в цитоплазму и либо используются самой клеткой, либо выводятся из нее. В цистернах комплекса Гольджи происходит синтез полисахаридов и их объединение с белками, в результате чего образуются гликопротеиды. Митохондрии — небольшие тельца палочковидной формы, ограниченные двумя мембранами. От внутренней мембраны митохондрии отходят многочисленные складки — кристы, на их стенках располагаются разнообразные ферменты, с помощью которых осуществляется синтез высокоэнергетического вещества — аденозинтрифосфорной кислоты (АТФ). В зависимости от активности клетки и внешних воздействий митохондрии могут перемещаться, изменять свои размеры, форму. В митохондриях найдены рибосомы, фосфолипиды, РНК и ДНК. С присутствием ДНК в митохондриях связывают способность этих органоидов к размножению путем образования перетяжки или почкованием в период деления клетки, а также синтез части митохондриальных белков. Лизосомы - мелкие овальные образования, ограниченные мембраной и рассеянные по всей цитоплазме. Встречаются во всех клетках животных и растений. Они возникают в расширениях эндоплазматической сети и в комплексе Гольджи, здесь заполняются гидролитическими ферментами, а затем обособляются и поступают в цитоплазму. В обычных" условиях лизосомы переваривают частицы, попадающие в клетку путем фагоцитоза, и органоиды отмирающих клеток. Продукты лизиса выводятся через мембрану лизосомы в цитоплазму, где они включаются в состав новых молекул. При разрыве лизоеомной мембраны ферменты поступают в цитоплазму и переваривают ее содержимое, вызывая гибель клетки. Пластиды есть только в растительных клетках и встречаются, у большинства зеленых растений. В пластидах синтезируются и накапливаются органические вещества. Различают пластиды трех видов: хлоропласты, хромопласты и лейкопласты. Хлоропласты — зеленые пластиды, содержащие зеленый пигмент хлорофилл. Они находятся в листьях, молодых стеблях, незрелых плодах. Хлоропласты окружены двойной мембраной. У высших растений внутренняя часть хлоропластов заполнена полужидким веществом, в котором параллельно друг другу уложены пластинки. Парные мембраны пластинок, сливаясь, образуют стопки, содержащие хлорофилл. В каждой стопке хлоропластов высших растений чередуются слои молекул белка и молекул липидов, а между ними располагаются молекулы хлорофилла. Такая слоистая структура обеспечивает максимум свободных поверхностей и облегчает захват и перенос энергии в процессе фотосинтеза. Хромопласты — пластиды, в которых содержатся растительные пигменты (красный или бурый, желтый, оранжевый). Они сосредоточены в цитоплазме клеток цветков, стеблей, плодов, листьев растений и придают им соответствующую окраску. Хромопласты образуются из лейкопластов или хлоропластов в результате накопления пигментов каротиноидов. Лейкопласты—бесцветные пластиды, располагающиеся в неокрашенных частях растений: в стеблях, корнях, луковицах и др. В лейкопластах одних клеток накапливаются зерна крахмала, в лейкопластах других клеток — масла, белки. Все пластиды возникают из своих предшественников — пропластид. В них выявлена ДНК, которая контролирует размножение этих органоидов. Клеточный центр, или центросома, играет важную роль при делении, клетки и состоит из двух центриолей. Он встречается у всех клеток животных и растений, кроме цветковых, низших грибов и некоторых, простейших. Центриоли в делящихся клетках принимают участие в формировании веретена деления и располагаются на его полюсах. В делящейся клетке первым делится клеточный центр, одновременно образуется ахроматиновое веретено, ориентирующее хромосомы при расхождении их к полюсам. В дочерние клетки отходит по одной центриоле. У многих растительных и животных клеток имеются органоиды специального назначения: реснички, выполняющие функцию движения (инфузории, клетки дыхательных путей), жгутики (простейшие одноклеточные, мужские половые клетки у животных и растений и др.). Включения - временные элемеаты, возникающие в клетке на определенной стадии ее жизнедеятельности в результате синтетической функции. Они либо используются, либо выводятся из клетки. Включениями являются также запасные питательные вещества: в растительных клетках—крахмал, капельки жира, блки, эфирные масла, многие органические кислоты, соли органических и неорганических кислот; в животных клетках - гликоген (в клетках печени и мышцах), капли жира (в подкожной клетчатке); Некоторые включения накапливаются в клетках как отбросы — в виде кристаллов, пигментов и др. Вакуоли — это полости, ограниченные мембраной; хорошо выражены в клетках растений и имеются у простейших. Возникают в разных участках расширений эндоплазматической сети. И постепенно отделяются от нее. Вакуоли поддерживают тургорное давление, в них сосредоточен клеточный или вакуолярный сок, молекулы которого определяют его осмотическую концентрацию. Считается, что первоначальные продукты синтеза - растворимые углеводы, белки, пектины и др. — накапливаются в цистернах эндоплазматической сети. Эти скопления и представляют собой зачатки будущих вакуолей. Цитоскелет. Одной из отличительных особенностей эукариотической клетки является развитие в ее цитоплазме скелетных образований в виде микротрубочек и пучков белковых волокон. Элементы цитоскелета тесно связаны с наружной цитоплазматической мембраной и ядерной оболочкой, образуют сложные переплетения в цитоплазме. Опорные элемеиты цитоплазмы определяют форму клетки, обеспечивают движение внутриклеточных структур и перемещение всей клетки. Ядро клетки играет основную роль в ее жизнедеятельности, с его удалением клетка прекращает свои функции и гибнет. В большинстве животных клеток одно ядро, но встречаются и многоядерные клетки (печень и мышцы человека, грибы, инфузории, зеленые водоросли). Эритроциты млекопитающих развиваются из клеток-предшественников, содержащих ядро, но зрелые эритроциты утрачивают его и живут недолго. Ядро окружено двойной мембраной, пронизанной порами, посредством которых оно тесно связано с каналами эндоплазматической сети и цитоплазмой. Внутри ядра находится хроматин — спирализованные участки хромосом. В период деления клетки они превращаются в палочковидные структуры, хорошо различимые в световой микроскоп. Хромосомы — это сложный комплекс белков с ДНК, называемый нуклеопротеидом. Функции ядра состоят в регуляции всех жизненных отправлений клетки, которую оно осуществляет при помощи ДНК и РНК-материальных носителей наследственной информации. В ходе подготовки к делению клетки ДНК удваивается, в процессе митоза хромосомы расходятся и передаются дочерним клеткам, обеспечивая преемственность наследственной информации у каждого вида организмов. Кариоплазма — жидкая фаза ядра, в которой в растворенном виде находятся продукты жизнедеятельности ядерных структур. Ядрышко — обособленная, наиболее плотная часть ядра. В состав ядрышка входят сложные белки и РНК, свободные или связанные фосфаты калия, магния, кальция, железа, цинка, а также рибосомы. Ядрышко исчезает перед началом деления клетки и вновь формируется в последней фазе деления. Таким образом, клетка обладает тонкой и весьма сложной организацией. Обширная сеть цитоплазматических мембран и мембранный принцип строения органоидов позволяют разграничить множество одновременно протекающих в клетке химических реакций. Каждое из внутриклеточных образований имеет свою структуру и специфическую функцию, но только при их взаимодействии возможна гармоничная жизнедеятельность клетки.На основе такого взаимодействия вещества из окружающей среды поступают в клетку, а отработанные продукты выводятся из нее во внешнюю среду — так совершается обмен веществ. Совершенство структурной организации клетки могло возникнуть только в результате длительной биологической эволюции, в процессе которой выполняемые ею функции постепенно усложнялись. Простейшие одноклеточные формы представляют собой и клетку, и организм со всеми его жизненными проявлениями. В многоклеточных организмах клетки образуют однородные группы — ткани. В свою очередь ткани формируют органы, системы, и их функции определяются общей жизнедеятельностью целостного организма. 2. Прокариотическая клетка. Помимо организмов с типичной клеточной организацией {эукариотические клетки) существуют относительно простые, доядерные, или прокариотические, клетки — бактерии и синезеленые, у которых отсутствуют оформленное ядро, окруженное ядерной мембраной, и высокоспециализированные внутриклеточные органоиды. Особую форму организации живого представляют вирусы и бактериофаги (фаги). Их строение крайне упрощено: они состоят из ДНК (либо РНК) и белкового футляра. Свои функции обмена веществ и размножения вирусы и фаги осуществляют только внутри клеток другого организма: вирусы — внутри клеток растений и животных, фаги - в бактериальных клетках как паразиты на, генетическом уровне. К прокариотам относят бактерии и сине-зелёные водоросли (цианеи). Наследственный аппарат прокариот представлен одной кольцевой молекулой ДНК, не образующей связей с белками и содержащей по одной копии каждого гена — гаплоидные организмы. В цитоплазме имеется большое количество мелких рибосом; отсутствуют или слабо выражены внутренние мембраны. Ферменты пластического обмена расположены диффузно. Аппарат Гольджи представлен отдельными пузырьками. Ферментные системы энергетического обмена упорядоченно расположены на внутренней поверхности наружной цитоплазматической мембраны. Снаружи клетка окружена толстой клеточной стенкой. Многие прокариоты способны к спорообразованию в неблагоприятных условиях существования; при этом выделяется небольшой участок цитоплазмы содержащий ДНК, и окружается толстой многослойной капсулой. Процессы метаболизма внутри споры практически прекращаются. Попадая в благоприятные условия, спора преобразуется в активную клеточную форму. Размножение прокариот происходит простым делением надвое. Средняя величина прокариотических клеток 5 мкм. У них нет никаких внутренних мембран, кроме впячиваний плазматической мембраны. Пласты отсутствуют. Вместо клеточного ядра имеется его эквивалент (нуклеоид), лишенный оболочки и состоящий из одной-единственной молекулы ДНК. Кроме того бактерии могут содержать ДНК в форме крошечных плазмид, сходных с внеядерными ДНК эукариот. В прокариотических клетках, способных к фотосинтезу (сине-зеленые водоросли, зеленые и пурпурные бактерии) имеются различно структурированные крупные впячивания мембраны – тилакоиды, по своей функции соответствующие пластидам эукариот. Эти же тилакоиды или – в бесцветных клетках – более мелкие впячивания мембраны (а иногда даже сама плазматическая мембрана) в функциональном отношении заменяют митохондрии. Другие, сложно дифференцированные впячивания мембраны называют мезасомами; их функция не ясна. Только некоторые органеллы прокариотической клетки гомологичны соответствующим органеллам эукариот. Для прокариот характерно наличие муреинового мешка – механически прочного элемента клеточной стенки Сравнительная характеристика клеток растений, животных, бактерий, грибов При сравнении бактерий с эукариотами можно выделить единственное сходство - наличие клеточной стенки, а вот сходства и различия эукариотических организмов заслуживают более пристального внимания. Следует начать сравнение с компонентов, которые свойственны и растениям, и животным, и грибам. Это ядро, митохондрии, Аппарат (комплекс) Гольджи, эндоплазматический ретикулум (или эндоплазматическая сеть) и лизосомы. Они характерны для всех организмов, имеют сходное строение и выполняют одинаковые функции. Теперь следует акцентировать внимание на различиях. Растительная клетка, в отличие от животной, имеет клеточную стенку, состоящую из целлюлозы. Кроме того, существую органеллы свойственные растительным клеткам - пластиды и вакуоли. Наличие этих компонентов обусловлено необходимостью растений поддерживать форму, при отсутствии скелета. Есть отличия и в особенностях роста. У растений он происходит в основном за счет увеличения размера вакуолей и растяжения клеток, в то время как у животных происходит увеличение объема цитоплазмы, а вакуоль вовсе отсутствует. Пластиды (хлоропласты, лейкопласты, хромопласты) характерны преимущественно для растений, поскольку их основная задача - это обеспечить автотрофный способ питания. У животных в противовес растениям существуют пищеварительные вакуоли, которые обеспечивают гетеротрофный способ питания. Грибы занимают особое положение и для их клеток характерны признаки свойственные и для растений, и для животных. Подобно животным грибам присущ гетеротрофный тип питания, содержащая хитин клеточная оболочка, а основным запасающим веществом является гликоген. В то же время для них, как для растений, характерен неограниченный рост, неспособность к передвижению и питание путем всасывания. |
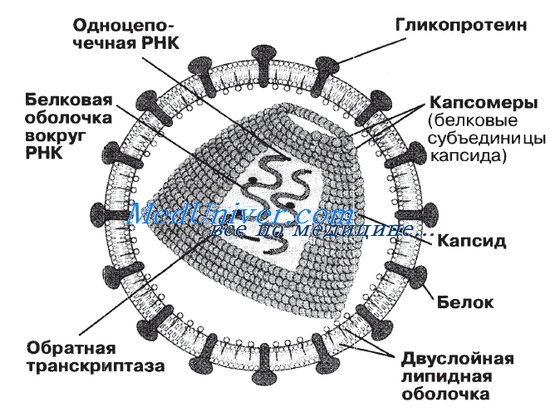
Строение вирусов
Полноценная по строению и инфекционная, т.е. способная вызвать заражение, вирусная частица вне клетки называется вирионом. Сердцевина («ядро») вириона содержит одну молекулу, а иногда две или несколько молекул нуклеиновой кислоты. Белковый чехол, покрывающий нуклеиновую кислоту вириона и защищающий ее от вредных воздействий окружающей среды, называется капсидом. Нуклеиновая кислота вириона является генетическим материалом вируса (его геномом) и представлена дезоксирибонуклеиновой кислотой (ДНК) или рибонуклеиновой кислотой (РНК), но никогда двумя этими соединениями сразу. (Хламидии, риккетсии и все другие «истинно живые» микроорганизмы содержат одновременно ДНК и РНК.) Нуклеиновые кислоты самых мелких вирусов содержат три или четыре гена, тогда как самые крупные вирусы имеют до ста генов.
У некоторых вирусов в дополнение к капсиду имеется еще и внешняя оболочка, состоящая из белков и липидов. Она образуется из мембран зараженной клетки, содержащих встроенные вирусные белки. Термины «голые вирионы» и «лишенные оболочки вирионы» используются как синонимы. Капсиды самых мелких и просто устроенных вирусов могут состоять лишь из одного или нескольких видов белковых молекул. Несколько молекул одного или разных белков объединяются в субъединицы, называемые капсомерами. Капсомеры, в свою очередь, образуют правильные геометрические структуры вирусного капсида. У разных вирусов форма капсида является характерной особенностью (признаком) вириона.
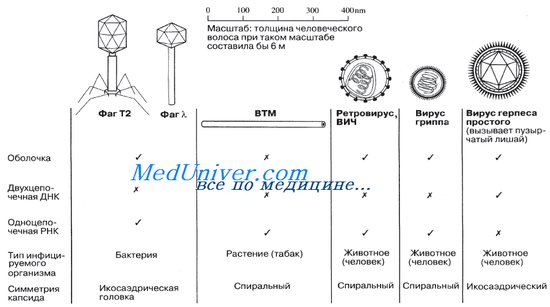
Вирионы со спиральным типом симметрии, как у вируса табачной мозаики, имеют форму удлиненного цилиндра; внутри белкового чехла, состоящего из отдельных субъединиц – капсомеров, находится свернутая спираль нуклеиновой кислоты (РНК). Вирионы с икосаэдрическим типом симметрии (от греч. eikosi – двадцать, hedra – поверхность), как у полиовируса, имеют сферическую, а точнее, многогранную форму; их капсиды построены из 20 правильных треугольных фасеток (поверхностей) и похожи на геодезический купол.
У отдельных бактериофагов (вирусов бактерий; фагов) смешанный тип симметрии. У т.н. «хвостатых» фагов головка имеет вид сферического капсида; от нее отходит длинный трубчатый отросток – «хвост».
Встречаются вирусы с еще более сложным строением. Вирионы поксвирусов (вирусы группы оспы) не имеют правильного, типичного капсида: между сердцевиной и наружной оболочкой у них располагаются трубчатые и мембранные структуры.
Основные структурные компоненты клетки
20 Августа 2009
Клетка является структурной и функциональной единицей живых организмов. Многие клетки человеческого организма имеют общее строение: они состоят из ядра и цитоплазмы, отделенных друг от друга и от окружающей среды мембранами. Цитоплазма содержит ряд органелл, различного рода включения, цитоскелет (промежуточные филаменты, микротрубочки, микрофиламенты). Клетка ограничена снаружи плазматической мембраной плазмолеммой.
Основные компоненты мембраны – белки и липиды, в небольшом количестве имеются углеводы и полисахариды. Мембрана представляет собой мозаичную структуру, основой которой является фосфолипидный бислой. Белки мембраны интегрированы в фосфолипиды с внутренней или внешней поверхности, образуя непрерывную структуру мембраны. В целом мембрана находится в жидком состоянии, т.е. белки и липиды свободно в ней перемешаются. Основной характеристикой биологических мембран является их полупроницаемость – избирательный перенос молекул различных веществ внутрь клетки и выведение их из неё. Проникновение низкомолекулярных веществ в клетку может осуществляться простой диффузией через липидный слой, высокомолекулярных путем эндоцитоза. Этот перенос осуществляется путем вворачивания плазмоллемы внутрь клетки и отшнуровки от нее мелких секреторных пузырьков. Обратный путь выведения веществ из клетки называется экзоцитоз. На наружной поверхности плазмолеммы имеются специализированные структуры рецепторы (гликопроленды), которые служат для взаимодействия с клетками и их медиаторами. В целом гликопротеиды образуют сплошной рыхлый слой гликокаликс, состав которого специфичен для каждого типа клеток, что играет важную роль в процессах распознавания и межклеточного взаимодействия.
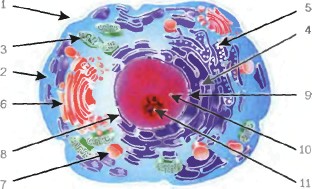 Схема
строения клетки: 1. плазмолемма; 2.
цитозоль; 3. митохондрии; 4. гладкая
эндоплазмотическая сеть; 5. шероховатая
эндоплазмотическая сеть; 6. аппарат
Гольджи; 7. лизосома; 8. кариолемма; 9. поры
ядерной мембраны; 10. кариоплазма; 11.
ядрышко.
Ядро наиболее важная структура
клетки, в нем сосредоточена основная
масса ДНК, являющаяся носителем
генетической информации. Большинство
клеток имеет одно ядро, однако встречаются
дву- и многоядерные клетки (плазматические,
остеокласты, мегакариоциты и др.) Ядро
ограничено мембраной (кариолеммой),
состоящей из двух липопротеидных слоев.
Наружная мембрана связана с рибосомами,
а к внутренней тесно прилежит хроматин
кариоплазмы. Наружная и внутренняя
мембраны сливаются в области ядерных
пор, через которые осуществляется
транспорт белков и РНК. Поры ядерной
мембраны заполнены белковым конгломератом,
который изолирует кариоплазму от
цитоплазмы, поэтому состав кариоплазмы,
в том числе по содержанию ионов, отличается
от состава цитоплазмы. Количество пор
увеличивается в клетках, активно
синтезирующих РНК.
Содержимое ядра
разделяют на хроматин, нуклеины (ядрышки)
и кариоплазму. Хроматин это нуклеопротеид,
представленный комплексом ДНК и белков.
В состав хроматина входит также
незначительное количество РНК. В ядре
фибриллярный компонент хроматина может
быть расположен рыхло, формируя видимый
в световом микроскопе диффузный хроматин.
Частично фибриллы хроматина могут быть
собраны вместе, образуя конденсированный
хроматин. Характер распределения
хроматина является важным признаком,
по которому дифференцируют различные
типы клеток, а также определяют их
степень зрелости.
Диффузный хроматин
характерен для молодых интенсивно
пролиферирующих клеток, по мере созревания
клетки он становится конденсированным.
В ядре осуществляется синтез ДНК и всех
видов РНК. Форма ядер поддерживается
преимущественно хроматином. Она может
быть разнообразной – округлая,
округло-овальная, продолговатая,
почковидная, сегментированная,
полиморфная, что и отличает различные
виды клеток. В нейтрофилах могут
встречаться дополнительные сегменты
(тельца Барра или половой хроматин),
представляющие собой конденсированную
Х-хромосому. Патология ядер клетки
приводит к нарушению ее синтетической
функции. Неестественная гибель
ядросодержащей клетки (аноптоз)
сопровождается фрагментацией ядра
(кариорекенс), превращением его в плотную
гомогенную массу (пикноз). Возможны
дегенеративные изменения ядер
(кариолизис).
Ядрышко (нуклеола) –
активно функционирующий в период
интерфазы локус хромосомы. В его состав
входит рибосомальная РНК, белки и
небольшое количество ДНК, в нем
сосредоточен важнейший фермент синтеза
РНК – полимераза. Общее число ядрышек
варьирует от 1 до 4-5 и больше. В ядрышке
образуются субъединицы рибосом (большая
и малая), которые затем выходят в
цитоплазму, где идет их сборка. Ядрышки
в клетках хорошо различимы в световом
микроскопе. Они окружены плотным
хроматином. Ядрышко может перемешаться
в пределах ядра и в состоянии активного
синтеза белка чаще находится вблизи
кариолеммы.
В ряде случаев нуклеолы
приходится дифференцировать от явления
хроматолиза. Хроматолиз – результат
разрушения ДНК эндонуклеазами.
Морфологически он представлен в виде
одной или нескольких четко очерченных
образований округлой или овальной
формы, розового цвета. Хроматолиз
является морфологическим признаком
поздних этапов аноптоза.
Все
пространство клетки, кроме ядра, относится
к цитоплазме которая включает в себя
гиалоплазму и расположенные в ней
основные органеллы клетки. Гиалоплазма
является внутренней средой клепки, в
которой осуществляются процессы обмена
и поддерживается клеточный гомеостаз.
В ее состав входят вода, белки, липиды.
ферменты, нуклеиновые кислоты,
неорганические и другие вещества. В
гиалоплазме расположены внутриклеточные
структуры. Она имеет хорошо развитую
структуру цитоскелета, в состав которого
входят микротрубочки, промежуточные
филаменты и микрофиламенты, участвующие
в движении внутриклеточных структур,
поддержании формы клетки. Цвет цитоплазмы
в зависимости от содержания в ней РНК
варьирует от интенсивно синего,
голубоватого до розового. В клетках с
выраженной белоксинтезирующей функцией
(плазматические клетки, эритробласты)
цитоплазма базофильного цвета.
Митохондрии
имеют вид тонких палочек, нитей, так же
как и ядро, имеют двойную мембрану.
Наружная часть – гладкая мембрана,
внутренняя образуем многочисленные
складки (крипты). Митохондрии имеют
собственную ДНК и рибосомную
белоксинтезирующую систему, которая
располагается в матриксе. Они способны
синтезировать лишь белки своего матрикса
и несколько белков, входящих в состав
внутренней мембраны. Митохондрии служат
энергетической станцией клетки, в них
осуществляются процессы окисления и
синтез АТФ. Помимо этого, они играют
важную роль в запуске аноптоза. Это
связано с тем, что митохондрии являются
источником цитохрома С, аноптоз-индуцирующего
фактора компонентов, необходимых для
передачи аноптотического сигнала в
ядро
клетки. Митохондрии лабильный
компонент цитоплазмы. Уменьшение их
числа или размеров наблюдается в к
клетках печени при диабете, голодании,
после облучения. Большое число
фрагментированных митохондрий, как
правило, характерно для состояния
повышенного энергообразования.
Эндоплазматическая
сеть система внутриклеточных канальцев,
вакуолей и цистерн. В клетках имеется
две разновидности эндоплазматической
сети – гладкая, лишенная рибосом, и
шероховатая, с прикрепленными к ней
рибосомами. Основная функция
эндоплазматической сети формирование
мембран и разграничение внутреннего
пространства клетки. Благодаря этому
в цитоплазме одновременно могут протекать
противоположно направленные процессы,
например синтез и распад глюкозы.
Большинство метаболических процессов
в клекте обеспечивается мембраносвязанными
ферментами, кофакторами, субстратами.
Эпдоплазматическая сеть является с
этой точки зрения, основой, на которой
функционируют внутриклеточные
метаболические процессы. В гладкой
эндоплазматической сети синтезируются
некоторые липиды, происходит нейтрализация
токсических веществ, образование гранул
гликогена. Хорошо развитая эндоплазматическая
сеть исключается в секретирующих клетках
(плазмоциты).
Рибосомы – гранулы,
которые свободно лежат в цитоплазме
либо прикреплены к кариолемме или
мембранам эндоплазматической сети.
Основная функция рибосом – синтез
белка. Поскольку в состав рибосом входит
много РНК, их скопления в цитоплазме
обеспечивают ее базофильную окраску.
Рибосомы, прикрепленные к эндоплазматической
сети, синтезируют белки, которые идут
на построение ее мембран; рибосомы,
свободно лежащие в цитоплазме, синтезируют
цитоплазматические белки, а прикрепленные
к кариолемме – ядерные белки.
Аппарат
Гольджи (пластинчатый комплекс) –
система специализированных мембран,
связанных с процессами секреции,
образованием клеточных мембран и
дизосом. Этот комплекс располагается
в перинуклеарной зоне и особенно развит
в секреторных клетках, что отличает их
наличием выраженной зоны просветления
вокруг ядра (плазматические клетки)
Лизосомы представляют собой мелкие
пузырьки, ограниченные однослойной
мембраной и содержащие внутри набор
гидролитических ферментов. Лизосомы –
производные аппарата Гольджи, с ними
связаны процессы внутриклеточного
расщепления белков, нуклеиновых кислот,
полисахаридов и др. При повреждении
лизосом или повышении проницаемости
их мембран наблюдается аутолиз клетки.
Гипертрофия лизосомного аппарата на
фоне воспалительных процессов и
интоксикации сопровождается появлением
токсогенной зернистости в цитоплазме
клетки.
Для многих клеток характерно
наличие гранул, цвет, размер и число
которых является важной характеристикой
идентификации клетки. В цитоплазме
возможно появление включений: тельца
Расселa (образованы за счет конденсации
парапротеина), тельца Ауэра (палочковидной
формы, состоят из агрегированных
азурофильных гранул), тельца Деле
(цитоплазмические включения, состоящие
из остатков цитоплазматической сети,
содержащей рибосомы), включения и
вакуолизация в цитоплазме нейрофилов
при синдроме Чедиака-Хигаси, вакуолизация
в бластах при лимфоме Беркитта. Некоторые
включения в цитоплазме представлены в
виде фагоцитированных ядер, эритроцитов
и тромбоцитов (гемофагоцитов).
Схема
строения клетки: 1. плазмолемма; 2.
цитозоль; 3. митохондрии; 4. гладкая
эндоплазмотическая сеть; 5. шероховатая
эндоплазмотическая сеть; 6. аппарат
Гольджи; 7. лизосома; 8. кариолемма; 9. поры
ядерной мембраны; 10. кариоплазма; 11.
ядрышко.
Ядро наиболее важная структура
клетки, в нем сосредоточена основная
масса ДНК, являющаяся носителем
генетической информации. Большинство
клеток имеет одно ядро, однако встречаются
дву- и многоядерные клетки (плазматические,
остеокласты, мегакариоциты и др.) Ядро
ограничено мембраной (кариолеммой),
состоящей из двух липопротеидных слоев.
Наружная мембрана связана с рибосомами,
а к внутренней тесно прилежит хроматин
кариоплазмы. Наружная и внутренняя
мембраны сливаются в области ядерных
пор, через которые осуществляется
транспорт белков и РНК. Поры ядерной
мембраны заполнены белковым конгломератом,
который изолирует кариоплазму от
цитоплазмы, поэтому состав кариоплазмы,
в том числе по содержанию ионов, отличается
от состава цитоплазмы. Количество пор
увеличивается в клетках, активно
синтезирующих РНК.
Содержимое ядра
разделяют на хроматин, нуклеины (ядрышки)
и кариоплазму. Хроматин это нуклеопротеид,
представленный комплексом ДНК и белков.
В состав хроматина входит также
незначительное количество РНК. В ядре
фибриллярный компонент хроматина может
быть расположен рыхло, формируя видимый
в световом микроскопе диффузный хроматин.
Частично фибриллы хроматина могут быть
собраны вместе, образуя конденсированный
хроматин. Характер распределения
хроматина является важным признаком,
по которому дифференцируют различные
типы клеток, а также определяют их
степень зрелости.
Диффузный хроматин
характерен для молодых интенсивно
пролиферирующих клеток, по мере созревания
клетки он становится конденсированным.
В ядре осуществляется синтез ДНК и всех
видов РНК. Форма ядер поддерживается
преимущественно хроматином. Она может
быть разнообразной – округлая,
округло-овальная, продолговатая,
почковидная, сегментированная,
полиморфная, что и отличает различные
виды клеток. В нейтрофилах могут
встречаться дополнительные сегменты
(тельца Барра или половой хроматин),
представляющие собой конденсированную
Х-хромосому. Патология ядер клетки
приводит к нарушению ее синтетической
функции. Неестественная гибель
ядросодержащей клетки (аноптоз)
сопровождается фрагментацией ядра
(кариорекенс), превращением его в плотную
гомогенную массу (пикноз). Возможны
дегенеративные изменения ядер
(кариолизис).
Ядрышко (нуклеола) –
активно функционирующий в период
интерфазы локус хромосомы. В его состав
входит рибосомальная РНК, белки и
небольшое количество ДНК, в нем
сосредоточен важнейший фермент синтеза
РНК – полимераза. Общее число ядрышек
варьирует от 1 до 4-5 и больше. В ядрышке
образуются субъединицы рибосом (большая
и малая), которые затем выходят в
цитоплазму, где идет их сборка. Ядрышки
в клетках хорошо различимы в световом
микроскопе. Они окружены плотным
хроматином. Ядрышко может перемешаться
в пределах ядра и в состоянии активного
синтеза белка чаще находится вблизи
кариолеммы.
В ряде случаев нуклеолы
приходится дифференцировать от явления
хроматолиза. Хроматолиз – результат
разрушения ДНК эндонуклеазами.
Морфологически он представлен в виде
одной или нескольких четко очерченных
образований округлой или овальной
формы, розового цвета. Хроматолиз
является морфологическим признаком
поздних этапов аноптоза.
Все
пространство клетки, кроме ядра, относится
к цитоплазме которая включает в себя
гиалоплазму и расположенные в ней
основные органеллы клетки. Гиалоплазма
является внутренней средой клепки, в
которой осуществляются процессы обмена
и поддерживается клеточный гомеостаз.
В ее состав входят вода, белки, липиды.
ферменты, нуклеиновые кислоты,
неорганические и другие вещества. В
гиалоплазме расположены внутриклеточные
структуры. Она имеет хорошо развитую
структуру цитоскелета, в состав которого
входят микротрубочки, промежуточные
филаменты и микрофиламенты, участвующие
в движении внутриклеточных структур,
поддержании формы клетки. Цвет цитоплазмы
в зависимости от содержания в ней РНК
варьирует от интенсивно синего,
голубоватого до розового. В клетках с
выраженной белоксинтезирующей функцией
(плазматические клетки, эритробласты)
цитоплазма базофильного цвета.
Митохондрии
имеют вид тонких палочек, нитей, так же
как и ядро, имеют двойную мембрану.
Наружная часть – гладкая мембрана,
внутренняя образуем многочисленные
складки (крипты). Митохондрии имеют
собственную ДНК и рибосомную
белоксинтезирующую систему, которая
располагается в матриксе. Они способны
синтезировать лишь белки своего матрикса
и несколько белков, входящих в состав
внутренней мембраны. Митохондрии служат
энергетической станцией клетки, в них
осуществляются процессы окисления и
синтез АТФ. Помимо этого, они играют
важную роль в запуске аноптоза. Это
связано с тем, что митохондрии являются
источником цитохрома С, аноптоз-индуцирующего
фактора компонентов, необходимых для
передачи аноптотического сигнала в
ядро
клетки. Митохондрии лабильный
компонент цитоплазмы. Уменьшение их
числа или размеров наблюдается в к
клетках печени при диабете, голодании,
после облучения. Большое число
фрагментированных митохондрий, как
правило, характерно для состояния
повышенного энергообразования.
Эндоплазматическая
сеть система внутриклеточных канальцев,
вакуолей и цистерн. В клетках имеется
две разновидности эндоплазматической
сети – гладкая, лишенная рибосом, и
шероховатая, с прикрепленными к ней
рибосомами. Основная функция
эндоплазматической сети формирование
мембран и разграничение внутреннего
пространства клетки. Благодаря этому
в цитоплазме одновременно могут протекать
противоположно направленные процессы,
например синтез и распад глюкозы.
Большинство метаболических процессов
в клекте обеспечивается мембраносвязанными
ферментами, кофакторами, субстратами.
Эпдоплазматическая сеть является с
этой точки зрения, основой, на которой
функционируют внутриклеточные
метаболические процессы. В гладкой
эндоплазматической сети синтезируются
некоторые липиды, происходит нейтрализация
токсических веществ, образование гранул
гликогена. Хорошо развитая эндоплазматическая
сеть исключается в секретирующих клетках
(плазмоциты).
Рибосомы – гранулы,
которые свободно лежат в цитоплазме
либо прикреплены к кариолемме или
мембранам эндоплазматической сети.
Основная функция рибосом – синтез
белка. Поскольку в состав рибосом входит
много РНК, их скопления в цитоплазме
обеспечивают ее базофильную окраску.
Рибосомы, прикрепленные к эндоплазматической
сети, синтезируют белки, которые идут
на построение ее мембран; рибосомы,
свободно лежащие в цитоплазме, синтезируют
цитоплазматические белки, а прикрепленные
к кариолемме – ядерные белки.
Аппарат
Гольджи (пластинчатый комплекс) –
система специализированных мембран,
связанных с процессами секреции,
образованием клеточных мембран и
дизосом. Этот комплекс располагается
в перинуклеарной зоне и особенно развит
в секреторных клетках, что отличает их
наличием выраженной зоны просветления
вокруг ядра (плазматические клетки)
Лизосомы представляют собой мелкие
пузырьки, ограниченные однослойной
мембраной и содержащие внутри набор
гидролитических ферментов. Лизосомы –
производные аппарата Гольджи, с ними
связаны процессы внутриклеточного
расщепления белков, нуклеиновых кислот,
полисахаридов и др. При повреждении
лизосом или повышении проницаемости
их мембран наблюдается аутолиз клетки.
Гипертрофия лизосомного аппарата на
фоне воспалительных процессов и
интоксикации сопровождается появлением
токсогенной зернистости в цитоплазме
клетки.
Для многих клеток характерно
наличие гранул, цвет, размер и число
которых является важной характеристикой
идентификации клетки. В цитоплазме
возможно появление включений: тельца
Расселa (образованы за счет конденсации
парапротеина), тельца Ауэра (палочковидной
формы, состоят из агрегированных
азурофильных гранул), тельца Деле
(цитоплазмические включения, состоящие
из остатков цитоплазматической сети,
содержащей рибосомы), включения и
вакуолизация в цитоплазме нейрофилов
при синдроме Чедиака-Хигаси, вакуолизация
в бластах при лимфоме Беркитта. Некоторые
включения в цитоплазме представлены в
виде фагоцитированных ядер, эритроцитов
и тромбоцитов (гемофагоцитов).