

Тема 3. Исследование постсинаптических потенциалов Введение
Высокоспециализированные функциональные контакты, существующие между нейроном и другими клетками (нервными, мышечными, железистыми), получили название синапсов. Исследования химической синаптической передачи были начаты в 20-30-е гг. XX в., в работах отечественных и зарубежных физиологов – А.Ф.Самойлова, Х.С.Коштоянца, А.В.Кибякова, О.Леви, Г.Дейла, В.Фельдберга, Дж.Экклса, и др.. Механизмы функционирования синапсов продолжают активно изучаться и в настоящее время. Классическими объектами для исследования химических синапсов являются нервно-мышечные соединения скелетных мышц позвоночных – концевые пластинки.
Миелинизированный аксон мотонейрона, вступая в контакт со скелетным мышечным волокном, постепенно утоньшается и теряет свою миелиновую оболочку. Поблизости от места формирования синаптического контакта аксон образует на поверхности волокна конечные разветвления, которые называются пресинаптическими нервными окончаниями или нервными терминалями.
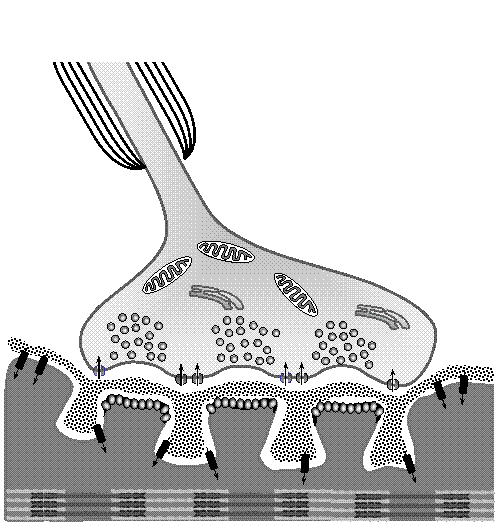
Мембрана мышечного волокна, находящаяся непосредственно под нервными терминалями, называется постсинаптической. У скелетных мышечных волокон высших позвоночных постсинаптическая мембрана и прилежащая к ней саркоплазма заметно выступают над общей поверхностью волокна, образуя как бы небольшой вырост, который назвали "подошвой", или "платформой" для контакта с нервным волокном, откуда и берет название этот тип синапсов - "концевая пластинка" (end platte). Постсинаптическая мембрана мышечного волокна имеет множество складок, расположенных непосредственно под пресинаптическими нервными окончаниями. В аксоплазме нервных терминалей с помощью электронной микроскопии обнаружено большое скопление характерных пузырьков - везикул. Медиатор нервно-мышечных синапсов – ацетилхолин (АХ) – содержится как в самой аксоплазме терминалей, так и в синаптических пузырьках. Однако его концентрация в пузырьках в 25-30 раз выше. Согласно классическим представлениям, под воздействием нервного импульса, распространяющегося от тела нейрона до его нервных терминалей, происходит деполяризация мембраны нервной терминали и высвобождение медиатора из везикул в синаптическую щель путем экзоцитоза.
Потенциал действия распространяется по аксону вплоть до последнего миелинизированного сегмента с постоянной амплитудой и скоростью. В области конечных разветвлений аксона амплитуда и скорость ПД могут снижаться в связи с тем, что здесь аксон утоньшается и разветвляется, и это замедляет распространение ПД. Деполяризация мембраны нервной терминали приводит к открыванию потенциал-активируемых натриевых, калиевых и кальциевых каналов на ее мембране. Кальциевые каналы сосредоточены в непосредственной близости к месту скопления пузырьков в примембранном пространстве нервной терминали, в так называемых “активных зонах”.
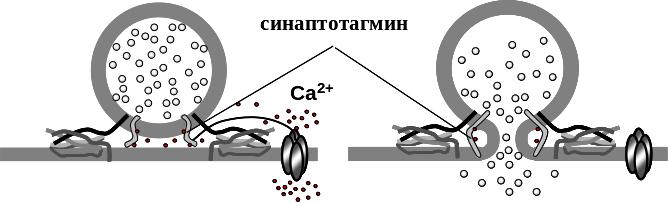
Часть везикул с помощью сложного комплекса специальных белков-сцепок – синаптотагмина, синаптобревина, синаптофизина и других – прикреплена к мембране терминали вблизи от кальциевых каналов и готова к экзоцитозу – высвобождению медиатора. Момент слияния везикулы с мембраной терминалии и выброс из нее медиатора происходит лишь в ответ на генерацию на мембране пресинаптического ПД, который приводит к открыванию потенцил-активируемых Са-каналов терминали и входу по ним ионов кальция. Это приводит к кратковременному повышению концентрации свободного ионизированного кальция вблизи примембранных синаптических пузырьков. Это, в свою очередь, ведет к слиянию везикул с пресинаптической мембраной и высвобождению из них ацетилхолина в синаптическую щель.
Молекулы ацетилхолина пересекают синаптическую щель и взаимодействуют с холинорецепторами, встроенными в постсинаптическую мембрану мышечного волокна. В результате этого взаимодействия изменяется конформация молекулы холинорецептора, являющейся одновременно канальным белком, и это приводит к открыванию ионного канала в молекуле холинорецептора. Ионный канал холинорецептора обладает относительно низкой ионоселективностью и проницаем и для ионов натрия, и для ионов калия, и отчасти – для ионов кальция, однако проницаемость для ионов натрия в 1,5-2 раза выше, чем для ионов калия, поэтому преимущественный ионный ток создают ионы натрия. Вход ионов натрия по каналам холинорецепторов приводит к кратковременной деполяризации постсинаптической мембраны, называемой постсинаптическим потенциалом, или потенциалом концевой пластинки (ПКП).
Электрофизиологический анализ амплитудно-временных параметров постсинаптических потенциалов привел исследователей к заключению, что молекулы ацетилхолина поступают в синаптическую щель не путем непрерывной диффузии, а дискретными порциями, или "квантами". Носителями таких порций-“квантов” АХ являются синаптические везикулы. Ах попадает в везикулу из аксоплазмы, где он синтезируется, с помощью специальной молекулы-транспортера, встроенной в мембрану везикулы. В результате в везикуле накапливается определенное количество медиатора, которое высвобождается в сианптическую щель при экзоцитозе везикулы и носит название «кванта» медиатора.
Установлено, один квант медиатора содержит около 10 000 молекул АХ. При высвобождении такой порции медиатора в синаптическую щель молекулы АХ диффундируют к постсинаптической мембране и взаимодействуют с молекулами холинорецепторов, расположенных на постсинаптической мембране. Электрофизиологические и авторадиографические исследования показали, что в результате высвобождения одного кванта АХ на постсинаптической мембране открывается 2000-2500 ионных каналов холинорецепторов, расположенных на площади порядка 2мкм2. Возникает входящий ионный ток и происходит кратковременная деполяризация постсинаптической мембраны; эти процессы можно наблюдать при внутриклеточном отведении потенциалов в виде отдельного миниатюрного потенциала концевой пластинки – минПКП. Итак, электрофизиологическим проявлением секреции АХ в виде "квантов" являются миниатюрные потенциалы концевой пластинки (минПКП). Они были зарегистрированы впервые в 1952 г. П.Фэттом и Б.Катцем в концевой пластинке портняжной мышцы лягушки. МинПКП возникали спонтанно, без внешних воздействий на нервное волокно; их амплитуда колебалась от 0,1 до 1,0мВ.
Случайный нерегулярный выброс отдельных квантов медиатора может происходить спонтанно, в отсутствии нервного импульса. Величины минПКП, регистрируемых в одном нервно-мышечном синапсе, примерно одинаковы и, несмотря на некоторые вариации амплитуд минПКП, их амплитудное распределение обычно соответствует нормальному гауссовому распределению. Этот факт свидетельствует об относительном постоянстве размеров квантов медиатора.
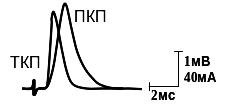
Временные параметры минПКП. На рис. видно, что отдельный ПКП достигает максимума в течение 1-1,5мс, а его спад носит экспоненциальный характер с постоянной времени 4-6мс. В случае генерации минПКП ионный ток, входящий через каналы холинорецепторов и текущий через Rсин (сопротивление каналов холинорецепторов для входящего синаптического тока) и Cм (емкость мембраны постсинаптической клетки) быстро перезаряжает (деполяризует) ее, а уже через 1-1,2мс каналы холинорецепторов закрываются, и ток прекращается. Поэтому период нарастания минПКП – это время, в течение которого входящий синаптический ток (минТКП) течет через rсин и Cм мембраны, а спад потенциала (минПКП) – функция постоянной времени мембраны (то есть отражает временной ход разрядки емкости мембраны Cм через пассивное сопротивление постсинаптической мембраны Rм). Таким образом, постоянная спада минПКП зависит от м=Rм.Cм (рис. 5.).
А. Б.
Рис. 5. А. Сравнение тока концевой пластинки (ТКП), зарегистрированного методом фиксации потенциала, и потенциала концевой пластинки (ПКП), зарегистрированного внутриклеточным микроэлектродом.
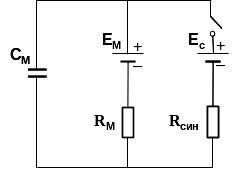
Б. Эквивалентная электрическая схема постсинаптической мембраны;
См - емкость постсинаптической мембраны,
Rм - пассивное сопротивление постсинаптической мембраны,
Rсин - сопротивление каналов холинорецепторов для входящего синаптического тока,
Eм - мембранный потенциал покоя, Ес - равновесный потенциал для входящего синаптического ионного тока.
Средняя амплитуда минПКП, регистрируемых в мышечном волокне диафрагмы мыши – порядка 1,0мВ. Амплитуда минПКП зависит (помимо размеров самого кванта медиатора) от уровня мембранного потенциала мышечного волокна, от сопротивления постсинаптической мембраны, а также от активности ацетилхолинэстеразы в синаптической щели.
Частота минПКП, т.е. число квантов, высвобождаемых в единицу времени из нервного окончания, – функция состояния пресинаптической нервной терминали. В отличие от амплитуды минПКП, их частота не зависит от состояния постсинаптической мембраны. Экспериментально установлено, что при деполяризации нервной терминали электрическим полем или под действием растворов с повышенным содержанием ионов калия частота минПКП увеличивается. При гиперполяризации терминали частота минПКП, напротив, снижается. В мышечных волокнах диафрагмы мыши частота регистрируемых минПКП, в среднем – около 1Гц.
Потенциалы концевой пластинки (ПКП), вызванные нервным импульсом. Как говорилось выше, высвобождение медиатора может происходить как спонтанно, так и под действием нервного импульса, приходящего к нервной терминали. Деполяризация нервной терминали вследствие генерации пресинаптического ПД на мембране, приводит к открыванию потенциал-зависимых Са2+-каналов на мембране терминали. Входящий по этим каналам кальций связывается с белками, встроенными в мембрану везикулы, что приводит к слиянию везикулы с мембраной терминали и экзоцитозу содержимого везикулы в синаптическую щель. В ответ на 1 пресинаптический ПД может одновременно подвергаться экзоцитозу от десятка до сотен везикул. Высвобождающийся из множества везикул медиатор действует на постсинаптическую мембрану и вызывает генерацию высокоамплитудного постсинаптического потенциала, называемого потенциалом концевой пластинки (ПКП). ПКП, возникающий в ответ на нервный импульс, является результатом одновременного действия на постсинаптическую мембрану 50-200 квантов медиатора. Амплитуда ПКП может колебаться от импульса к импульсу и достигать величин 20-50мВ. Колебания амплитуды ПКП связаны с тем, что число везикул, готовых к экзоцитозу в каждый данный момент времени в ответ на действие на них ионов кальция, непостоянно.
Число
квантов, высвобождаемых из терминали
в ответ на нервный импульс, можно
ограничить путем уменьшения концентрации
ионов кальция и повышения концентрации
ионов магния в наружной среде. При этом
становится хорошо заметно, что флуктуации
амплитуды ПКП всегда кратны величине
одного кванта медиатора.
Можно подсчитать квантовый состав (m)
одного ПКП путем деления средней
амплитуды ПКП на среднюю амплитуду
минПКП:
![]()
![]()
Таким образом, эффективность синаптической передачи регулируется путем изменения числа выделяемых квантов медиатора. Изучение эффектов одиночных квантов медиатора (регистрируемых в виде минПКП) и эффектов, вызываемых одновременным высвобождением многих квантов АХ (регистрируемых в виде ПКП), обеспечивает возможность детального анализа пре- и постсинаптических механизмов функционирования химического синапса.
Влияние ингибиторов ацетпилхолинэстеразы. Ацетилхолинэстераза (АХЭ) – фермент, расщепляющий молекулы АХ на холин и уксусную кислоту. Молекулы АХЭ находятся в синаптическй щели: они встроены в базальную мембрану, покрывающую постсинаптическую мембрану. Особый интерес представляет действие ингибиторов АХЭ (армина, прозерина и др.). Ингибиторы резко увеличивают продолжительность действия ацетилхолина, выделяемого из нервных окончаний. При этом возрастает длительность синаптических потенциалов и их амплитуда. Эксперименты показали, что несмотря на присутствие в синаптической щели АХЭ все кванты АХ (хотя и в редуцированном виде) достигают постсинаптической мембраны. Назначение АХЭ сводится лишь к тому, чтобы ограничить время пребывания медиатора в синаптической щели вблизи холинорецепторов. В нормальных условиях ацетилхолинэстераза, быстро гидролизуя молекулы АХ, укорачивает время присутствия медиатора в щели, а также обеспечивает быстрое и независимое действие отдельных квантов АХ, ограничивая зону распространения молекул ацетилхолина, входящих в один квант.
В условиях инактивации АХЭ с помощью ингибиторов появляется возможность повторных связываний негидролизованных молекул ацетилхолина с рецепторами постсинаптической мембраны. Происходит, по-видимому, частичное перекрытие зон действия отдельных квантов медиатора, в результате повторные взаимодействия АХ с ХР продлевают временной ход синаптического тока, увеличивая время нарастания и спада минПКП.