

Типы местностей и урочищ
Тип местности |
Тип урочищ |
Пойменная местность (Рис.2а) |
затапливаемая пойм а (Рис.2б); истинная пойма (Рис.2в) |
Склоновая местность (Рис.2д) |
семейство овражно-балочных урочищ (Рис.2г); верхний крутой склон (Рис.2е); средний пологий склон (Рис.2ж); нижний пологий склон (Рис.2з) |
Возвышенная водораздельная местность (Рис.2и) |
водораздельная слабодренированная территория (Рис.2к) (равнинная (Рис.2л), холмистая (Рис.2м)); водораздельная дренируемая территория (Рис.2н); останцовые горы (Рис.2о) |
Выделенные нами типы местностей и урочищ в пределах бассейна р. Чардым представлены на рис.2(а,б,в,г,д,е,ж,з,и,к,л,м,н,о)
а |
б |
в |
г |
д |
е |
ж |
з |
и |
к |
л |
м |
н |
о |
|
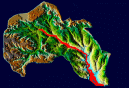













Рис. 2. Типы местностей и урочищ в пределах бассейна р. Чардым
Полевые исследования в бассейне р. Чардым позволили скорректировать границы выделенных нами типов местности и урочищ и определить их площади.
Таблица 2
Площади типов местности и урочищ
Тип местности, площадь |
Тип урочищ, площадь |
Пойменная местность, 67,2 км² (Рис.2а) |
затапливаемая пойма;26,3 км² (Рис.2б) истинная пойма 40,9 км² (Рис.2в) |
Склоновая местность, 1054,2 км² (Рис.2д) |
семейство овражно-балочных урочищ 477,5 км²(Рис.2г) верхний крутой склон 146 км²; (Рис.2е) средний пологий склон 189,8 км²; (Рис.2ж) нижний пологий склон 240,9 км²; (Рис.2з) |
Возвышенная водораздельная местность, 338,7 км² (Рис.2и) |
водораздельная слабодренированная территория,109,5 км² (Рис.2к) (равнинная19 км² (Рис.2л), холмистая 90,5 км² (Рис.2м)); водораздельная дренируемая территория 195,6 км² (Рис.2н); останцовые горы 33,6 км² (Рис.2о) |
Анализ таблицы позволил выделить доминантные типы местностей. В нашем случае доминантным типом будет склоновая местность, которая занимает 72,2% от всего бассейна реки Чардым. Внутри склоновой местности наибольшую площадь занимают овражно-балочные урочища - 32,7% от всего бассейна. На основе этой карты также можно прогнозировать развитие экологических процессов, формировать программу природоохранных мероприятий, разрабатывать план природообустройства различных природно-территориальных комплексов, проводить качественную оценку земель.
Список использованных источников:
Программный комплекс GeoControl2 v2.1 Build 54 (Win32)
Влияние объема жизненного пространства на продуктивность гидробионтов в токсической среде
Могильная Е.C., Гандзюра В.П.
Киевский национальный университет имени Тараса Шевченко, Киев, Украина, katyam@bigmir.net
Продукционная гидробиология и водная токсикология развивались в XX веке как независимые и не связанные друг с другом научные направления. Между тем в антропогенно трансформированных водных экосистемах биопродукционные процессы зачастую протекают в условиях токсифицированной среды и существенно ею модифицируются [5].
Недоучет этого обстоятельства может приводить к существенным ошибкам в биопродукционных расчетах и оценке состояния гидроэкосистем по биопродукционным показателям. Анализ литературы по данному вопросу показал, что с одной стороны, исследование биологической продуктивности в подавляющем большинстве случаев проводится без учета состояния качества среды обитания, в частности, экотоксикологической ситуации [1-5, 9-10], а с другой – что имеющийся в литературе колоссальный материал по продуктивности гидробионтов, их популяций и сообществ практически не используется для оценки состояния водных экосистем. В то же время не раз отмечалась перспективность применения биопродукционных показателей для оценки экотоксикологической ситуации [1, 5-11, 17].
За последние годы вышло ряд работ, посвященных установлению зависимости между биопродукционными показателями разных видов гидробионтов и их популяций от уровня токсичности водной среды, что позволяет использовать разные биопродукционные показатели как интегральные критерии уровня токсического загрязнения водоемов [1, 5, 9-10].
Следует отметить, что биопродукционные показатели у разных видов гидробионтов имеют разную чувствительность к токсическому загрязнению, но в целом тенденция их изменений имеет общий характер, что позволяет использовать их для оценки уровня загрязнения водной среды (рис. 1

Рис. 1. Удельная скорость роста у Elodea canadensis (E.c.), Lemna trisulca (L.t.) и Pelmatohydra oligactis (P.o.) при разной концентрации Pb2+ в воде
Следовательно, биопродукционные показатели являются информативными интегральными критериями токсичности водной среды обитания. Установлены общие закономерности влияния токсического загрязнения среды на удельную скорость роста и накопления энергии, эффективность трансформации вещества и энергии, величину накопленной в биомассе энергии на единицу доступного ее потока, соотношение между величинами накопленной в биомассе энергии и уровнем дыхания [9]. Показано, что даже незначительное повышение уровня тяжелых металлов в воде (от 0,1 ПДК) вызывает резкие колебания значений всех биопродукционных показателей организмов и популяций, что позволяет проводить раннюю диагностику токсических эффектов. Выявлены существенные нарушения структуры энергетического баланса биосистем в токсической среде [10], особенности влияния величины доступной биосистеме энергии на уровень проявления токсических эффектов на разных стадиях токсикоза.
Однако при этом практически не изученным остался вопрос о влиянии объема жизненного пространства на все исследуемые показатели, что может чревато неадекватной оценкой экотоксикологической ситуации по биопродукционным показателям гидробионтов [14, 16].
В связи с этим целью нашей работы было установление взаимосвязи между объемом жизненного пространства, уровнем токсичности среды и темпом роста рыб, выяснение влияния этого параметра на их токсикорезистентность и адекватное проведение оценок состояния водных экосистем с учетом вышеупомянутых факторов.
Материал и методы исследования. Использовали общепринятые методы физиологических, ихтиологических [15], гидрохимических и гидроэкологических [13, 18] исследований. Результаты обработаны с использованием методов статистического анализа и стандартных компьютерным программ.
Эксперимпенты проводили на молоди золотой рыбки Carassius auratus auratus (Linnaeus, 1758); гуппи Poecilia reticulatа (Peters, 1859) и данио рерио Danio rerio Hamilton, 1822). Температуру воды поддерживали в пределах 20±10С с помощью терморегуляторов, аэрацию осуществляли с помощью микрокомпрессоров. Источником свинца служил Pb(NO3)2, никеля – NiSO4, хрома – K2Cr2O7. Каждые 5 суток рыб взвешивали. Кормили трубочником Tubifex tubifex. Интенсивность дыхания определяли в замкнутых респирометрах, концентрацию кислорода определяли по методу Винклера [18]. Удельную скорость роста (g) определяли про формуле g = (ln m2 – ln m1) / (t2 - t1) = ln (m2/m1)/∆t [12]. Валовую эффективность использования пищи на рост (К1) определяли как отношение приростов массы тела к массе использованной за это время рациона [12].
Полученные результаты и их обсуждение. Результаты экспериментов с молодью рыб показали, что незначительное увеличение содержания ионов ТМ в воде вызывает существенные колебания значений биопродукционных показателей. Во всех случаях при повышенных концентрациях ТМ в воде резко возрастала дисперсия значений биопродукционных показателей и среднее отклонение, причем максимальных значений она достигает при концентрации хрома 0,0005 мг/л.
Средние отклонения значений удельной скорости роста на порядок превышали аналогичные величины в контроле. Исследование структуры энергетического баланса гидробионтов в токсической среде позволило установить резкое возрастание энерготрат на поддержание жизнедеятельности, причем наиболее существенно возрастает стандартный обмен.
Таким образом, нами было установлено, что в условиях токсической среды резко возрастают энергетические траты на поддержание жизнедеятельности гидробионтов.
Следующим этапом наших исследований было установление оптимального объема жизненного пространства для рыб. Эксперименты с Данио рерио показали, что интенсивность дыхания непосредственно зависит от величины жизненного пространства. Причем максимальная интенсивность дыхания была у одиночных особей, а с увеличением числа особей до 5 экз/10 л она закономерно снижалась, и при плотности 5-10 экз/л была почти на одном уровне. При плотности посадки, близкой к 100 экз./л она снова возрастала (табл. 1).
Таблица 1.