

Структура и функции нейрона
Структурной единицей нервной системы является нервная клетка, или нейрон. Нейроны отличаются от других клеток организма многими ' особенностями. Прежде всего их популяция, насчитывающая от 10 до 30 млрд. (а быть может, и больше1) клеток, почти полностью «укомплектована» уже к моменту рождения, и ни один из нейронов, если он отомрет, не замещается новым. Принято считать, что после того, как человек минует период зрелости, у него ежедневно отмирает около 10 тысяч нейронов, а после 40 лет этот суточный показатель удваивается.
Другая особенность нейронов состоит в том, что в отличие от клеток других типов они ничего не продуцируют, не секретируют и не структурируют; единственная их функция заключается в проведении нервной информации.
Структура нейрона
Существует много типов нейронов, структура которых варьирует в зависимости от выполняемых ими в нервной системе функций; сенсорный нейрон отличается по своему строению от моторного нейрона или нейрона мозговой коры (рис. А.28).
Но какой бы ни была функция нейрона, все нейроны состоят из трех основных частей: тела клетки, дендритов и аксона.
Тело нейрона, как и всякой другой клетки, состоит из цитоплазмы и ядра. Цитоплазма нейрона, однако, особенно богата митохондриями, ответственными за выработку энергии, необходимой для поддержания высокой активности клетки. Как уже отмечалось, скопления тел нейронов образуют нервные центры в виде ганглия, в котором число клеточных тел исчисляется тысячами, ядра, где их еще больше, или, наконец, коры, состоящей из миллиардов нейронов. Тела нейронов образуют так называемое серое вещество.
Дендриты служат нейрону своего рода антеннами. Некоторые нейроны имеют много сотен дендритов, принимающих информацию от рецепторов или других нейронов и проводящих ее к телу клетки и ее единственному отростку другого типа-аксону.
Аксон представляет собой часть нейрона, ответственную за передачу информации дендритам других нейронов, мышцам или железам. У одних нейронов длина аксона достигает метра, у других аксон очень короткий. Как правило, аксон ветвится, образуя так называемое терминальное дерево; на конце каждой ветви имеется синоптическая бляшка. Именно она и образует соединение (синапс) данного нейрона с дендри-тами или телами других нейронов.
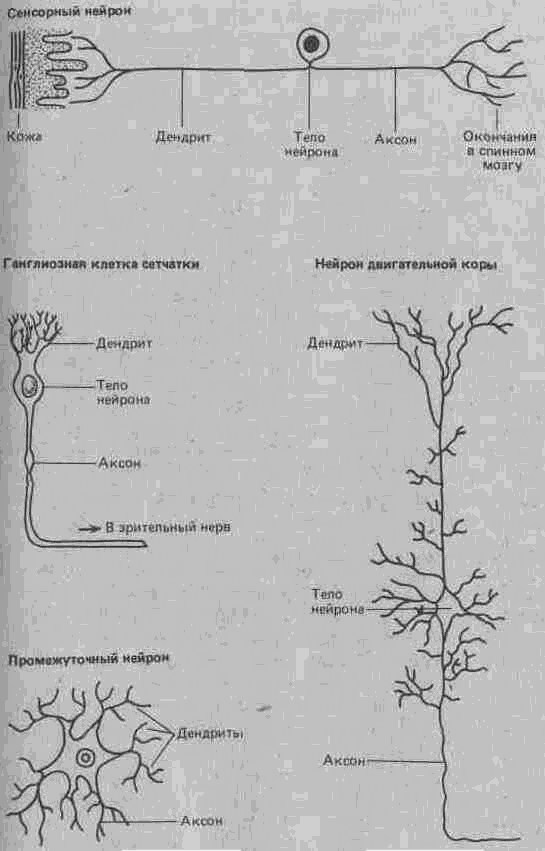
Рис. А.28. Различные типы нейронов.
Большинство нервных волокон (аксонов) покрыто оболочкой, состоящей из миелина - белого жироподобного вещества, выполняющего функции изоляционного материала. Миелиновая оболочка с регулярными промежутками в 1-2 мм прерывается перетяжками - перехватами Ранвье, которые увеличивают скорость пробегания нервного импульса по волокну, позволяя ему «перепрыгивать» с одного перехвата на другой, вместо того чтобы постепенно распространяться вдоль волокна. Сотни и тысячи собранных в пучки аксонов образуют нервные пути, которые благодаря миелину имеют вид белого вещества.
Нервный импульс
Информация поступает в нервные центры, перерабатывается там и затем передается эффекторам в виде нервных импульсов, пробегающих по нейронам и соединяющим их нервным путям.
Независимо от того, какую информацию передают нервные импульсы, пробегающие по миллиардам нервных волокон, они ничем не отличаются друг от друга. Почему же в таком случае импульсы, идущие от уха, передают информацию о звуках, а импульсы от глаза - о форме или цвете предмета, а не о звуках или о чем-нибудь совсем ином? Да просто потому, что качественные различия между нервными сигналами определяются не самими этими сигналами, а тем местом, куда они приходят: если это мышца, она будет сокращаться или растягиваться;
если это железа, она будет выделять секрет, уменьшать или прекращать секрецию; если это определенная область мозга, в ней будет формироваться зрительный образ внешнего стимула или же сигнал подвергнется расшифровке в виде, например, звуков. Теоретически достаточно было бы изменить ход нервных путей, например, часть зрительного нерва в зону мозга, ответственную за расшифровку звуковых сигналов, чтобы заставить организм «слышать глазами».
Потенциал покоя и потенциал действия
Нервные импульсы передают по дендритам и аксонам не сам внешний стимул как таковой и даже не его энергию. Внешний стимул лишь активирует соответствующие рецепторы, и эта активация преобразуется в энергию электрического потенциала, который создается на кончиках дендритов, образующих контакты с рецептором.
Возникающий при этом нервный импульс можно грубо сравнить с огнем, бегущим вдоль бикфордова шнура и поджигающим расположенный у него на пути патрон с динамитом; «огонь», таким образом, распространяется по направлению к конечной цели за счет небольших следующих друг за другом взрывов. Передача нервного импульса, однако, принципиально отличается от этого тем, что почти сразу же после прохождения разряда потенциал нервного волокна восстанавливается.
Нервное волокно в состоянии покоя можно уподобить маленькой батарейке; с наружной стороны его мембраны имеется положительный заряд, а с внутренней-отрицательный (рис. А.29), и этот потенциал покоя преобразуется в электрический ток только при замыкании обоих полюсов. Именно это и происходит при прохождении нервного импульса, когда мембрана волокна на какое-то мгновение становится проницаемой и деполяризуется. Вслед за этой деполяризацией наступает период рефрактерности, в течение которого мембрана реполяризуется и восста навливает способность к проведению нового импульса1. Так за счет последовательных деполяризации и происходит распространение этого потенциала действия (т.е. нервного импульса) с постоянной скоростью, варьирующей в пределах от 0,5 до 120 метров в секунду в зависимости от типа волокна, его толщины и наличия или отсутствия у него миелиновой. оболочки.
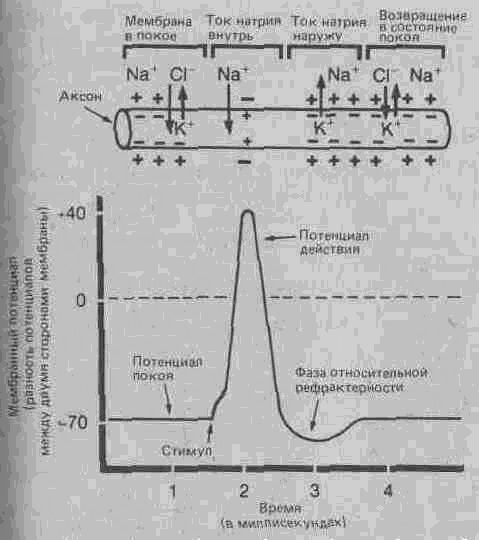
Рис. А.29. Потенциал действия. Развитие потенциала действия, сопровождающееся изменением электрического напряжения (от — 70 до + 40 мВ), обусловлено восстановлением равновесия между положительными и отрицательными ионами по обе стороны мембраны, проницаемость которой на короткое время увеличивается.
Закон «всё или ничего». Поскольку каждому нервному волокну присущ определенный электрический потенциал, распространяющиеся по нему импульсы независимо от интенсивности или "каких-либо других свойств внешнего стимула всегда имеют одни и те же характеристики. Это означает, что импульс в нейроне может возникнуть только в том случае, если его активация, вызванная стимуляцией рецептора или
импульсом от другого нейрона, будет превосходить некий порог, ниже которого активация неэффективна; но, если порог достигнут, сразу же возникает «полномерный» импульс. Этот факт получил название закона «всё или ничего».
Синаптическая передача
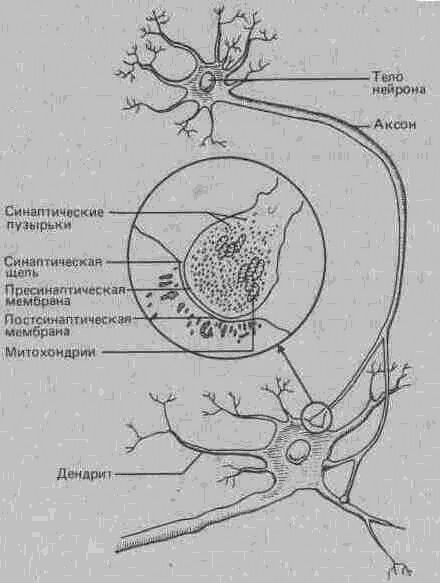
Рис. А.ЗО. Синаптическое соединение нейронов (в середине - область синапса при большем увеличении). Терминальная бляшка пресинаптического нейрона содержит пузырьки с запасом нейромедиатора и митохондрии, доставляющие энергию, необходимую для передачи нервного сигнала.
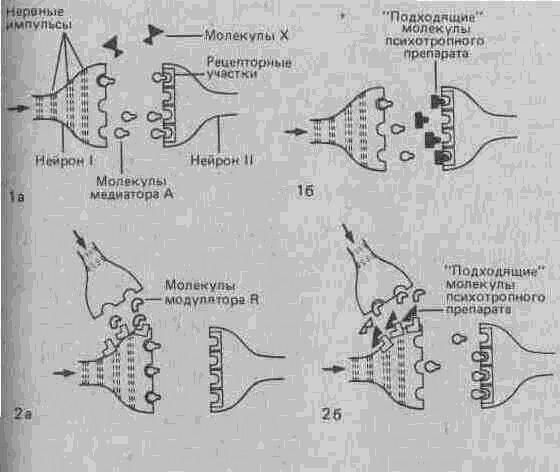
Рис. А.31. la. Медиатор А, молекулы которого освобождаются из концевой бляшки нейрона I, связывается специфическими рецепторами на дендритах нейрона II. Молекулы X, которые по своей конфигурации не подходят к этим рецепторам, занять их не могут и потому не вызывают каких-либо синаптических эффектов.
16. Молекулы М (например, молекулы некоторых психотропных препаратов) сходны по своей конфигурации с молекулами нейромедиатора А и поэтому могут связываться с рецепторами для этого медиатора, таким образом мешая ему выполнять свои функции. Например,. ЛСД мешает серотонину подавлять проведение сенсорных сигналов.
2а и 26. Некоторые вещества, называемые нейромодуляторами, способны воздействовать на окончание аксона, облегчая или подавляя высвобождение нейромедиатора.
Синапс. Синапсом называют область соединения между окончанием аксона одного нейрона и дендритами или телом другого. Каждый нейрон может образовать до 800-1000 синапсов с другими нервными клетками, а плотность этих контактов в сером веществе мозга составляет боле 600 млн. на 1 ми3 (рис. А.ЗО)1.
Место перехода нервного импульса с одного нейрона на другойпредставляет собой, собственно, не точку контакта, а скорее узкий промежуток, называемый синоптической щелью. Речь идет о щели шириной от 20 до 50 нанометров (миллионных долей миллиметра), которая с одной стороны ограничена мембраной пресинаптической бляшки нейрона, передающего импульс, а с другой-постсинаптической мембраной дендрита или тела другого нейрона, принимающего нервный сигнал и затем передающего его дальше.
Нейромедиаторы. Именно в синапсах происходят процессы, в результате которых химические вещества, освобождаемые пресинаптической мембраной, передают нервный сигнал с одного нейрона на другой. Эти вещества, получившие название нейромедиаторов (или просто медиаторов), своего рода «мозговые гормоны» (нейрогормоны) накапливаются в пузырьках синаптических бляшек и освобождаются, когда по аксону сюда приходит нервный импульс.
После этого медиаторы диффундируют в синаптическую щель и присоединяются к специфическим рецепторным участкам постсинаптической мембраны, т. е. к таким участкам, к которым они «подходят, как ключ к замку». В результате этого проницаемость постсинаптической мембраны изменяется, и таким образом сигнал передается с одного нейрона на другой; медиаторы могут также и блокировать передачу нервных сигналов на уровне синапса, уменьшая возбудимость постсинаптического нейрона.
Выполнив свою функцию, медиаторы расщепляются или нейтрализуются ферментами либо всасываются обратно в пресинаптическое окончание, что приводит к восстановлению их запаса в пузырьках к моменту прихода следующего импульса (рис. А.31).
Возбуждающая или тормозная функция синапса зависит главным образом от типа выделяемого им медиатора и от действия последнего на постсинаптическую мембрану. Некоторые медиаторы всегда оказывают только возбуждающее действие, другие - только тормозное (ингибирующее), а третьи в одних отделах нервной системы играют роль активаторов, а в других-ингибиторов.
Функции главных нейромедиаторов. В настоящее время известно несколько десятков этих нейрогормонов, но их функции изучены пока недостаточно. Сказанное, например, относится к ацетилхолину, который участвует в мышечном сокращении, вызывает замедление сердечного и дыхательного ритма и инактивируется ферментом ацетилхолинэстеразой1. Не вполне изучены и функции таких веществ из группы моноаминов, как норадреналин, отвечающий за бодрствование мозговой коры и учащение сердечного ритма, дофамин, присутствующий в «центрах удовольствия» лимбической системы и некоторых ядрах ретикулярной формации, где он участвует в процессах избирательного внимания, или серотонин, который регулирует сон и определяет объем информации, циркулирующей в сенсорных путях. Частичная инактивация моноаминов происходит в результате их окисления ферментом моноаминоксидазой. Этот процесс, обычно возвращающий активность мозга к нормальному уровню, в некоторых случаях может приводить к чрезмерному ее снижению, что в психологическом плане проявляется у человека в чувстве подавленности (депрессии).
Гамма-аминомасляная кислота (ГАМК) представляет собой нейро-медиатор, выполняющий примерно ту же физиологическую функцию, что и моноаминоксидаза. Ее действие состоит главным образом в снижении возбудимости мозговых нейронов по отношению к нервным импульсам.
Наряду с нейромедиаторами существует группа так называемых нейромодуляторов, которые в основном участвуют в регуляции нервного ответа, взаимодействуя с медиаторами и видоизменяя их эффекты. В качестве примера можно назвать вещество Р и брадикинин, участвующие в передаче болевых сигналов. Освобождение этих веществ в синапсах спинного мозга, однако, может быть подавлено секрецией эндорфи-нов и энкефалина, которая таким образом приводит к уменьшению потока болевых нервных импульсов (рис. А.31, 2а). Функции модуляторов выполняют и такие вещества, как фактор S, играющий, по-видимому, важную роль в процессах сна, холецистокинин, ответственный за чувство сытости, ангиотензин, регулирующий жажду, и другие агенты.
Нейромедиаторы и действие психотропных веществ. В настоящее время известно, что различные психотропные препараты действуют на уровне синапсов и тех процессов, в которых участвуют нейромедиаторы и нейромодуляторы.
Молекулы этих препаратов по своей структуре сходны с молекулами определенных медиаторов, что и позволяет им «обманывать»,различные механизмы синаптической передачи. Таким образом они нарушают действие истинных нейромедиаторов, либо занимая их место на рецепторных участках, либо мешая им всасываться обратно в пресинаптические окончания или подвергаться разрушению специфическими ферментами (рис. А.31, 26).
Установлено, например, что ЛСД, занимая серотониновые рецептор-ные участки, мешает серотонину затормаживать приток сенсорных сигналов. Таким образом ЛСД открывает доступ к сознанию для самых .разнообразных стимулов, непрерывно атакующих органы чувств.
Кокаин усиливает эффекты дофамина, занимая его место в рецептор-ных участках. Подобным же образом действуют морфин и другие опиаты, мгновенный эффект которых объясняется тем, что они быстро успевают занять рецепторные участки для эндорфинов1.
Действие амфетаминов обусловлено тем, что они подавляют обратное поглощение норадреналина пресинаптическими окончаниями. В результате накопление избыточного количества нейрогормона в синаптической щели приводит к чрезмерной степени бодрствования мозговой коры.
Принято считать, что эффекты так называемых транквилизаторов (например, валиума) объясняются главным образом их облегчающим влиянием на действие ГАМК в лимбической системе, что приводит к усилению - тормозных эффектов этого медиатора. Наоборот, как антидепрессанты действуют главным образом ферменты, инактивирую-Щие ГАМК, или такие препараты, как, например, ингибиторы моноамйноксидазы, введение которых увеличивает количества моноаминов в синапсах.
Смерть от некоторых отравляющих газов наступает вследствие удушья. Такое действие этих газов связано с тем, что их молекулы блокируют секрецию фермента, разрушающего ацетилхолин. Между тем ацетилхолин вызывает сокращение мышц и замедление сердечного и дыхательного ритма. Поэтому его накопление в синаптических пространствах приводит к угнетению, а затем и полной блокаде сердечной и дыхательной функций и одновременному повышению тонуса всей мускулатуры.
Изучение нейромедиаторов еще только начинается, и можно ожидать, что в скором времени будут открыты сотни, а может быть и тысячи этих веществ, многообразные функции которых определяют их первостепенную роль в регуляции поведения.