

Функции продолговатого мозга[править | править исходный текст]
Защитные рефлексы (например, кашель, чихание).
Жизненно важные рефлексы (например, дыхание).
Регулирование сосудистого тонуса.
Регулирование дыхательной системы
Рефлекторные центры продолговатого мозга:
пищеварение
сердечная деятельность
защитная (кашель, чихание и тому подобное)
центры регуляции тонуса скелетных мышц для поддержания позы человека.
укорочение или удлинение времени спинального рефлекса
11. Мозжечок, его строение и роль в регуляции движений (координацию движений). Рост и развитие мозжечка. Усложнение двигательных реакций ребенка в связи с развитием мозжечка. Последствия разрушения мозжечка.
Мозжечок (cerebellum; синоним малый мозг) — отдел головного мозга, обеспечивающий координацию движений, мышечный тонус и равновесие тела. Мозжечок расположен в задней черепной ямке над продолговатым мозгом и мостом. Над мозжечком находятся затылочные доли большого мозга (см. Головной мозг); между ними и мозжечком натянута палатка (или намет) мозжечка — отросток твердой мозговой оболочки. Анатомия и физиология. В М. различают верхнюю и нижнюю поверхности, передний и задний края. Мозжечок состоит из среднего отдела, или червя, и двух полушарий, каждое из которых разделяется бороздами на три доли (рис.). Каждой доле полушария соответствует определенный участок червя.
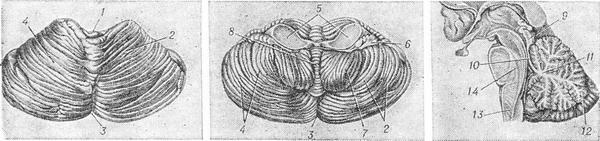 Мозжечок
(строение): а — верхняя, или задняя,
поверхность; б — нижняя, или передняя,
поверхность; в — сагиттальный разрез
через червь. 1 — передняя вырезка; 2 —
правое полушарие; 3 — задняя вырезка; 4
— левое полушарие; 5 — средние ножки
мозжечка: 6 — клочок; 7 — миндалина; 8 —
червь мозжечка; 9 — пластинка четверохолмия;
10 — передний мозговой парус; 11 — белое
вещество; 12 — кора мозжечка; 13 —
продолговатый мозг; 14 — мост.
Мозжечок
(строение): а — верхняя, или задняя,
поверхность; б — нижняя, или передняя,
поверхность; в — сагиттальный разрез
через червь. 1 — передняя вырезка; 2 —
правое полушарие; 3 — задняя вырезка; 4
— левое полушарие; 5 — средние ножки
мозжечка: 6 — клочок; 7 — миндалина; 8 —
червь мозжечка; 9 — пластинка четверохолмия;
10 — передний мозговой парус; 11 — белое
вещество; 12 — кора мозжечка; 13 —
продолговатый мозг; 14 — мост.
В мозжечке различают заложенное внутри белое вещество и покрывающее его тонким слоем серое корковое вещество. Белое вещество полушарий мозжечка соединяется медиально с белым веществом червя. Картина расположения белого вещества, видимая на сагиттальном разрезе червя, вследствие своего сходства с картиной разветвления дерева называется древом жизни. В белом веществе имеются еще скопления серого вещества — ядра М., из которых более важными являются зубчатые ядра крыши и ядра шатра. Белое вещество полушарий мозжечка соединяется с соседними частями головного мозга посредством волокнистых пучков. Эти пучки образуют различной толщины тяжи, называемые ножками мозжечка, и соединяют мозжечок с мостом, со средним и продолговатым мозгом. Средние ножки выходят из мозжечка латерально и, постепенно сближаясь, направляются вперед, переходя в мост. Верхние, или передние, ножки расположены медиально от средних, направляются вперед и в виде уплощенных круглых тяжей (также постепенно сходящихся) исчезают под четверохолмием, в области красных ядер. Между ними помещается передний мозговой парус. Нижние ножки идут назад и вниз к продолговатому мозгу. Главной функцией М. является регуляция согласованной (координированной) деятельности скелетных мышц. Вместе с корой головного мозга мозжечок участвует в координации так называемых произвольных движений. Осуществляется это благодаря связям мозжечка срецепторами, заложенными в скелетных мышцах, суставах и сухожилиях. Вместе с вестибулярным аппаратом полукружных каналов внутреннего уха (см.), сигнализирующим в центральную нервную систему о положении головы и тела в пространстве, мозжечок участвует в регуляции равновесия тела (см.) при ходьбе и активных движениях. Регуляция мозжечком координации движений скелетных мышц осуществляется через специальные системы проводящих волокон, идущих от мозжечку к передним рогам спинного мозга, где берут начало периферические двигательные нервы скелетных мышц. Патология. При поражении мозжечка развиваются главным образом расстройства согласованной деятельности скелетных мышц, а именно: нарушения координации произвольных движений и нарушения равновесия тела. Первая группа мозжечковых расстройств движений проявляется в нарушениях плавных движений конечностей (главным образом рук) и, в частности, в появлении дрожания (см.) в конце целенаправленного движения; в нарушениях речи (так называемая скандированная речь, в которой появляется не смысловая, а ритмическая расстановка ударений в словах); в замедленности произвольных движений и речи; в изменении почерка. Мозжечковые нарушения равновесия проявляются главным образом в головокружениях и изменении походки (см. Атаксия), которая приобретает характер походки пьяного человека, причем больной пошатывается в сторону очага поражения. Все эти расстройства иногда сопровождаются нистагмом (подергивание глазных яблок при их отведении). Частым симптомом поражения мозжечка является расстройство согласованной деятельности мышц, принадлежащих к разным мышечным группам, при их участии в одном моторном акте. Такая асинергия мышц ног и торса проявляется, например, при попытке больного без помощи рук принять сидячее положение из лежачего. Среди опухолей мозжечка чаще всего встречаются инфильтративно растущие доброкачественные новообразования, астроцитомы, ангиоретикулемы.
12. Промежуточный мозг. Таламус и его ядра. Функции таламуса. Гипоталамус и его роль.
Промежуточный мозг
Промежуточный мозг (diencephalon) интегрирует сенсорные, двигательные и вегетативные реакции, необходимые для целостной деятельности организма. Основными образованиями промежуточного мозга являются таламус, гипоталамус, который состоит из свода и эпифиза, и таламической области, которая включает в себя таламус, эпиталамус и метаталамус.
Таламус
Таламус (thalamus, зрительный бугор) — структура, в которой происходит обработка и интеграция практически всех сигналов, идущих в кору большого мозга от спинного, среднего мозга, мозжечка, базальных ганглиев головного мозга.
Морфофункциональная организация. В ядрах таламуса происходит переключение информации, поступающей от экстеро-, проприорецепторов и интероцепторов и начинаются таламокортикальные пути.
Учитывая, что коленчатые тела таламуса являются подкорковыми центрами зрения и слуха, а узел уздечки и переднее зрительное ядро участвуют в анализе обонятельных сигналов, можно утверждать, что зрительный бугор в целом является подкорковой «станцией» для всех видов чувствительности. Здесь раздражения внешней и внутренней среды интегрируются, после чего поступают в кору большого мозга.
Зрительный бугор является центром организации и реализации инстинктов, влечений, эмоций. Возможность получать информацию о состоянии множества систем организма позволяет таламусу участвовать в регуляции и определении функционального состояния организма в целом (подтверждением тому служит наличие в таламусе около 120 разнофункциональных ядер). Ядра образуют своеобразные комплексы, которые можно разделить по признаку проекции в кору на 3 группы: передняя проецирует аксоны своих нейронов в поясную извилину коры большого мозга; медиальная — в лобную долю коры; латеральная — в теменную, височную, затылочную доли коры. По проекциям определяется и функция ядер. Такое деление не абсолютно, так как одна часть волокон от ядер таламуса идет в строго ограниченные корковые образования, другая — в разные области коры большого мозга.
Ядра таламуса функционально по характеру входящих и выходящих из них путей делятся на специфические, неспецифические и ассоциативные.
К специфическим ядрам относятся переднее вентральное, медиальное, вентролатеральиое, постлатеральное, постмедиальное, латеральное и медиальное коленчатые тела. Последние относятся к подкорковым центрам зрения и слуха соответственно.
Основной функциональной единицей специфических таламических ядер являются «релейные» нейроны, у которых мало дендритов и длинный аксон; их функция заключается в переключении информации, идущей в кору большого мозга от кожных, мышечных и других рецепторов.
От специфических ядер информация о характере сенсорных стимулов поступает в строго определенные участки III—IV слоев коры большого мозга (соматотопическая локализация). Нарушение функции специфических ядер приводит к выпадению конкретных видов чувствительности, так как ядра таламуса, как и кора большого мозга, имеют соматотопическую локализацию. Отдельные нейроны специфических ядер таламуса возбуждаются рецепторами только своего типа. К специфическим ядрам таламуса идут сигналы от рецепторов кожи, глаз, уха, мышечной системы. Сюда же конвергируют сигналы от интерорецепторов зон проекции блуждающего и чревного нервов, гипоталамуса.
Латеральное коленчатое тело имеет прямые эфферентные связи с затылочной долей коры большого мозга и афферентные связи с сетчаткой глаза и с передними буграми четверохолмий. Нейроны латеральных коленчатых тел по-разному реагируют на цветовые раздражения, включение, выключение света, т. е. могут выполнять детекторную функцию.
В медиальное коленчатое тело (МТК) поступают афферентные импульсы из латеральной петли и от нижних бугров четверохолмий. Эфферетные пути от медиальных коленчатых тел идут в височную зону коры большого мозга, достигая там первичной слуховой области коры. МКТ имеет четкую тонотопичность. Следовательно, уже на уровне таламуса обеспечивается пространственное распределение чувствительности всех сенсорных систем организма, в том числе сенсорных посылок от интерорецепторов сосудов, органов брюшной, грудной полостей.
Ассоциативные ядра таламуса представлены передним медиодорсальным, латеральным дорсальным ядрами и подушкой. Переднее ядро связано с лимбической корой (поясной извилиной), медиодорсальное — с лобной долей коры, латеральное дорсальное — с теменной, подушка — с ассоциативными зонами теменной и височной долями коры большого мозга.
Основными клеточными структурами этих ядер являются мультиполярные, биполярные трехотростчатые нейроны, т. е. нейроны, способные выполнять полисенсорные функции. Ряд нейронов изменяет активность только при одновременном комплексном раздражении. На полисенсорных нейронах происходит конвергенция возбуждений разных модальностей, формируется интегрированный сигнал, который затем передается в ассоциативную кору мозга. Нейроны подушки связаны главным образом с ассоциативными зонами теменной и височной долей коры большого мозга, нейроны латерального ядра — с теменной, нейроны медиального ядра — с лобной долей коры большого мозга.
Неспецифические ядра таламуса представлены срединным центром, парацентральным ядром, центральным медиальным и латеральным, субмедиальным, вентральным передним, парафасцикулярным комплексами, ретикулярным ядром, перивентрикулярной и центральной серой массой. Нейроны этих ядер образуют свои связи по ретикулярному типу. Их аксоны поднимаются в кору большого мозга и контактируют со всеми ее слоями, образуя не локальные, а диффузные связи. К неспецифическим ядрам поступают связи из РФ ствола мозга, гипоталамуса, лимбической системы, базальных ганглиев, специфических ядер таламуса.
Возбуждение неспецифических ядер вызывает генерацию в коре специфической веретенообразной электрической активности, свидетельствующей о развитии сонного состояния. Нарушение функции неспецифических ядер затрудняет появление веретенообразной активности, т. е. развитие сонного состояния.
Сложное строение таламуса, наличие в нем взаимосвязанных специфических, неспецифических и ассоциативных ядер, позволяет ему организовывать такие двигательные реакции, как сосание, жевание, глотание, смех. Двигательные реакции интегрируются в таламусе с вегетативными процессами, обеспечивающими эти движения.
Конвергенцию сенсорных стимулов в таламус обусловливает возникновение так называемых таламических неукротимых болей, которые возникают при патологических процессах в самом таламусе.
13. Кора больших полушарий, доли, борозды, извилины. Нейронная организация коры, центры коры и их функции. Высший анализ раздражений в коре больших полушарий.
КОРА БОЛЬШИХ ПОЛУШАРИЙ ГОЛОВНОГО МОЗГА
(cortex hemispheria cerebri), паллиум, или плащ, слой серого вещества (1—5 мм), покрывающий полушария большого мозга млекопитающих. Эта часть головного мозга, развившаяся на поздних этапах эволюции, играет исключительно важную роль в осуществлении высшей нервной деятельности, участвует в регуляции и координации всех функций организма. У человека кора составляет примерно 44% объёма всего полушария, её поверхность в среднем 1468—1670 см2. В ходе эволюции сначала появляется древняя кора (палеокортекс) у рыб. С переходом животных к наземному существованию кора интенсивно развивается: у земноводных, кроме древней, намечается старая кора (архикортекс), у пресмыкающихся, кроме архи- и палеокортекса, появляются зачатки новой коры (неокортекс), к-рая достигает наибольшего развития у млекопитающих и особенно у человека. Поверхность неокортекса у человека занимает 95,6%, архикортекса 2,2%, палеокортекса 0,6%, промежуточной коры (отделяет неокортекс от палео- и архикортекса) 1,6% по отношению к поверхности полушария. Если представить кору мозга в виде единого покрова (плаща),одевающего поверхность полушарий, то осн. центр, часть его составит неокортекс, а древняя, старая и промежуточная кора займут место по краям этого плаща. Развитие коры в эволюции отражает осн. этапы совершенствования воспринимающей и интегрирующей деятельности мозга и управления целенаправленным двигат. поведением. У высших млекопитающих в связи с неравномерностью роста отд. структур неокортекса поверхность коры становится складчатой, покрытой бороздами и извилинами (гирэнцефалич. тип); у низших — поверхность коры гладкая (лиссэнцефалич. тип). Развивающаяся раньше других латеральная, или силь-виева, борозда отделяет височную долю от лобной и теменной. Выше и впереди сильвиевой борозды формируется поперечная центральная, или роландова, борозда, отделяющая лобную долю от теменной. Кроме этих основных борозд, большое число других отделяет друг от друга извилины коры. Борозды и извилины увеличивают поверхность коры без увеличения объёма черепа. Так, у человека ок. 2/3 поверхности всей коры расположены в глубине борозд. Строение коры характеризуется упорядоченностью с горизонтально-вертикальным распределением нейронов по слоям и колонкам. Структурно-функц. единица коры — модуль (объединение, блок), состоящий из пирамидных, звёздчатых и веретенообразных клеток, а также волокон, сосудов и клеток глии, и имеющий диам. ок. 100—150 мкм. Апикальные (верхушечные) дендриты пирамидных клеток и выходящие из коры их аксоны объединены в пучки. К модулям конвергирует множество разл. влияний (возбуждающих и тормозных). В результате их объединения (интеграция) посредством пространственно-временной суммации местных электрич. потенциалов на мембране клеток формируются синхронные импульсные залпы. Такие элементарные модули входят в более обширные объединения нейронов (колонки) с диам. до 1 мм. Др. структурным элементом коры является нейроглия, к-рая вместе с нейронами образует единый функциональный комплекс. Различия в строении отд. участков коры (плотность расположения, величина нейронов, их организация по слоям и колонкам) определяют архитектуру коры, или её цитоархитектонику. Кора имеет тесные связи с нижележащими структурами мозга, к-рые направляют к ней свои нервные волокна и сами находятся под контролем определ. корковых зон, получая от них по нервным путям регулирующие влияния. В составе коры выделяют проекционные (первичные и вторичные сенсорные), ассоциативные (третичные мультисенсорные) и интегративно-пусковые (моторные и др.) поля, что связано со сложным характером переработки информации и формирования программы целенаправленного поведения. В эволюции разл. функции организма оказываются всё более чётко представленными в коре мозга (кортиколизация функций). (см. ГОЛОВНОЙ МОЗГ, КОНЕЧНЫЙ МОЗГ).

Рнс. 1. Соотношение новой, древней, старой н промежуточной коры в головном мозге человека: 1 — большие полушария; 2 — мозжечок; 3 — продолговатый мозг; 4 — мозолистое тело; 5 — зрительные бугры. Горизонтальный штрих — новая кора, косой перекрёстный — древняя, вертикальный — старая, прямой перекрёстный — промежуточная.

Рис. 2. Поверхность коры головного мозга человека (вид сбоку): 1 — лобные извилины; 2 — центральная борозда; 3 — центральные извилины; 4 — теменные извилины; 5 — затылочные извилины; 6 — височные извилины; 7 — латеральная (сильвиева) борозда.
14. Условные и безусловные рефлексы (видовые). Неоднократное повторение сигнала и его подкрепление – условия образования условных рефлексов. Биологическое значение условных рефлексов.
Поведение человека связано с условно-безусловной рефлекторной деятельностью и представляет собой высшую нервную деятельность, результатом которой является изменение соотношения организма с внешней средой.
В отличие от высшей нервной деятельности низшая нервная деятельность состоит в совокупности реакций, направленных на объединение, интеграцию функций внутри организма.
Высшая нервная деятельность проявляется в виде сложных рефлекторных реакций, осуществляемых при обязательном участии коры больших полушарий и ближайших к ней подкорковых образований.
Впервые представление о рефлекторном характере деятельности головного мозга было широко и подробно развито основоположником отечественной физиологии И. М. Сеченовым в его книге "Рефлексы головного мозга". Идейная установка этого классического труда выражена в первоначальном, измененном под влиянием цензуры заглавии: "Попытка ввести физиологические основы в психические процессы". До И. М. Сеченова физиологи и неврологи не решались даже поставить вопрос о возможности объективного, чисто физиологического анализа психических процессов. Последние оставались полностью отданными на откуп субъективной психологии.
Идеи И. М. Сеченова получили блестящее развитие в замечательных трудах И. П. Павлова, открывшего пути объективного экспериментального исследования функций коры больших полушарий и создавшего стройное учение о высшей нервной деятельности.
И. П. Павлов показал, что в то время как в нижележащих отделах центральной нервной системы - подкорковых ядрах, мозговом стволе, спинном мозгу - рефлекторные реакции осуществляются по прирожденным, наследственно закрепленным нервным путям, в коре больших полушарий нервные связи вырабатываются и создаются в процессе индивидуальной жизни животных и человека, в результате сочетания бесчисленных, действующих на организм раздражений.
Открытие этого факта позволило разделить всю совокупность рефлекторных реакций, происходящих в организме, на две основные группы: на безусловные и условные рефлексы.
Различия условных и безусловных рефлексов |
Условные рефлексы
это реакции, приобретаемые организмом в процессе индивидуального развития на основе "жизненного опыта"
являются индивидуальными: у одних представителей одного и того же вида они могут быть, а у других отсутствуют
непостоянны и в зависимости от определенных условий они могут выработаться, закрепиться или исчезнуть; это их свойство и отражено в самом их названии
могут образоваться на самые разнообразные раздражения, приложенные к различным рецептивным полям
замыкаются на уровне коры. После удаления коры больших полушарий выработанные условные рефлексы исчезают и остаются только безусловные.
осуществляются через функциональные временные связи
Условные рефлексы вырабатываются на базе безусловных рефлексов. Для образования условного рефлекса необходимо сочетание времени какого-либо изменения внешней среды и внутреннего остояния организма, воспринятого корой больших полушарий, с осуществлением того или иного безусловного рефлекса. Только при этом условии изменение внешней среды или внутреннего состояния организма становится раздражителем условного рефлекса - условным раздражителем, или сигналом. Раздражение, вызывающее безусловный рефлекс, - безусловное раздражение - должно при образовании условного рефлекса сопутствовать условному раздражению, подкреплять его.
Для того чтобы звон ножей и вилок в столовой или стук чашки, из которой кормят собаку, вызывал выделение слюны в первом случае у человека, во втором случае у собаки, необходимо повторное совпадение этих звуков с едой - подкрепление вначале индифферентных в отношении слюнной секреции раздражителей кормлением, т. е. безусловным раздражением слюнных желез. Равным образом, вспыхивание электрической лампочки перед глазами собаки или звук звонка только в том случае вызовут условно-рефлекторное сгибание лапы, если они многократно сопровождались электрическим раздражением кожи ноги, вызывающим при всяком его применении безусловный сгибательный рефлекс. Подобно этому плач ребенка и отдергивание им ручек от горящей свечи будут наблюдаться только при условии, если предварительно вид свечи хотя бы один раз совпал с ощущением ожога. Во всех приведенных примерах являющиеся вначале относительно индифферентными внешние агенты - звон посуды, вид горящей свечи, вспыхивание электрической лампочки, звук звонка - становятся условными раздражителями в случае подкрепления их безусловными раздражениями. Лишь при этом условии индифферентные вначале сигналы внешнего мира становятся раздражителями определенного вида деятельности. |
Для образования условных рефлексов необходимо возникновение временной связи, замыкания между корковыми клетками, воспринимающими условное раздражение, и корковыми нейронами, входящими в состав дуги безусловного рефлекса.
При совпадении и сочетании условного и безусловного раздражений устанавливается связь между различными нейронами в коре полушарий мозга и между ними происходит процесс замыкания.
Безусловные рефлексы
это врожденные, наследственно передающиеся реакции организма
являются видовыми, т. е. свойственными всем представителям данного вида
относительно постоянны, как правило, сохраняются в течение всех жизни
осуществляются в ответ на адекватные раздражения, приложенные к одному определенному рецептивному полю
замыкаются на уровне спинного мозга и стволовой части головного мозга
осуществляются через филогенетически закрепленную, анатомически выраженную рефлекторную дугу.
Нужно отметить, однако, что у человека и обезьяны, у которых имеется высокая степень кортикализации функций, многие сложные безусловные рефлексы осуществляются при обязательном участии коры больших полушарий. Это доказывается тем, что ее поражения у приматов приводят к патологическим нарушениям безусловных рефлексов и исчезновению некоторых из них.
Следует также подчеркнуть, что далеко не все безусловные рефлексы появляются сразу к моменту рождения. Многие безусловные рефлексы, например, связанные с локомоцией, половым актом, возникают у человека и животного через длительный срок после рождения, но они обязательно появляются при условии нормального развития нервной системы.
Классификация условных и безусловных рефлексов |
Всю совокупность безусловных и образованных на их основе условных рефлексов принято по их функциональному значению делить на ряд групп.