

Молекулярные механизмы сокращения скелетной мышцы
Согласно теории скольжения нитей, мышечное сокращение происходит благодаря скользящему движению актиновых и миозиновых филламентов друг относительно друга. Механизм скольжения нитей включает несколько последовательных событий.
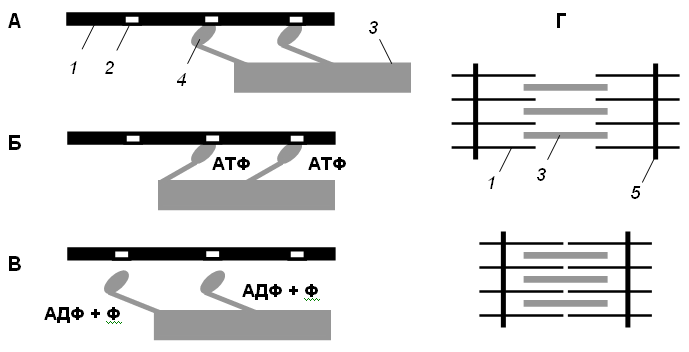
• Головки миозина присоединяются к центрам связывания актинового филламента (рис. 2, А).
• Взаимодействие миозина с актином приводит к конформационным перестройкам молекулы миозина. Головки приобретают АТФазную активность и поворачиваются на 120 ° . За счет поворота головок нити актина и миозина передвигаются на «один шаг» друг относительно друга (рис. 2, Б).
• Рассоединение актина и миозина и восстановление конформации головки происходит в результате присоединения к головке миозина молекулы АТФ и ее гидролиза в присутствии Са++ (рис. 2, В).
• Цикл «связывание – изменение конформации – рассоединение – восстановление конформации» происходит много раз, в результате чего актиновые и миозиновые филламенты смещаются друг относительно друга, Z -диски саркомеров сближаются и миофибрилла укорачивается
Расслабление скелетной мышцы
Расслабление мышцы вызывается обратным переносом ионов Са++ посредством кальциевого насоса в каналы саркоплазматического ретикулума. По мере удаления Са++ из цитоплазмы открытых центров связывания становится все меньше и в конце концов актиновые и миозиновые филламенты полностью рассоединяются; наступает расслабление мышцы.
Контрактурой называют стойкое длительное сокращение мышцы, сохраняющееся после прекращения действия раздражителя. Кратковременная контрактура может развиваться после тетанического сокращения в результате накопления в саркоплазме большого количества Са++ ; длительная (иногда необратимая) контрактура может возникать в результате отравления ядами, нарушений метаболизма.
20. Механизм проведения возбуждения по миелиновым и немиелизированным волокнам. Законы проведения возбуждения.
Механизм проведения возбуждения по нервному волокну.
Основная функция нервных волокон – передача нервного импульса. Нервные волокна представляют собой отростки нейронов. Существует 2 основных вида волокон: миелиновые и безмиелиновые.
Миелиновые нервные волокна, в отличие от безмиелиновых, покрыты одним или несколькими слоями клеток. Эта оболочка защищает волокно от повреждения, способствует его питанию, а также намного увеличивает скорость передачи нервного импульса. Через равные промежутки (в среднем через 1 мм) миелиновая оболочка прерывается, оставляя небольшие участки отростка нервной клетки – перехваты Ранвье.
Нервные волокна подразделяют на группы:
А – нервные волокна с самой толстой миелиновой оболочкой. Наиболее высокая скорость передачи нервного импульса.
В – миелиновая оболочка тоньше, скорость проведения возбуждения ниже
С – безмиелиновые волокна с относительно низкой скоростью передачи импульса.
При раздражении нервного волокна в его участке непосредственно соприкасающемся с раздражителем, возникает потенциал действия. Изменение заряда клеточной мембраны ведет к возникновению разности потенциалов между возбужденным и невозбужденным участками нервного волокна и следовательно к появлению электрического тока, направленного от возбужденного участка к невозбужденному.
В миелиновых волокнах импульс возникает только в перехватах Ранвье. При возбуждении каждый следующий перехват усиливает нервный импульс, а потому он не только не затухает в процессе передачи, но может «перепрыгивать через один или несколько перехватов. Это ведет к очень быстрому движению импульса по нервному волокну.
В безмиелиновых волокнах нервный импульс распространяется волнообразно, последовательно возбуждаются небольшие участки размером в несколько микрометров. Возбуждение распространяется гораздо медленнее, чем по миелиновым волокнам.
Нейрон как структурно – функциональная еденица ЦНС. Классификация нейронов, функциональные структуры нейрона. Механизм возникнвения возбуждения.
Нейрон – анатомо-гистологическая единица ЦНС. Он состоит из тела и отростков. Тела нейронов составляют серое вещество головного мозга. Их функции заключаются в переработке и хранении информации, а также в питании отростков.
Отростки нейронов:
аксоны – длинные маловетвистые отростки, проводящие информацию от тела нейрона к периферии
дендриты – короткие, сильноветвистые отростки, передающие информацию от периферии к телу нейрона.
Функции отростков заключаются в проведении информации к телу и от тела нейрона, в обеспечении взаимодействия нейронов с другими структурами.
По локализации нейроны подразделяются на центральные и периферические. Центральными называются те нейроны, тела которых лежат в пределах ЦНС. Периферические нейроны принадлежат периферической нервной системе. Они могут залегать в спинно-мозговых ганглиях, в ганглиях черепно-мозговых нервов, в ганглиях вегетативной нервной системы.
В зависимости от выполняемой функции нейроны делятся на 3 основные группы:
афферентные (чувствительные)
эфферентные (двигательные)
вставочные (контактные).
Афферентные нейроны обеспечивают восприятие раздражения и передачу информации в ЦНС.
Эфферентные нейроны обеспечивают передачу информации от ЦНС на периферию.
Вставочные нейроны обеспечивают передачу информации внутри ЦНС (с афферентных нейронов на эфферентные).
В зависимости от эффекта вставочные нейроны подразделяются на:
возбуждающие – оказывают возбуждающее влияние на эфферентные нейроны
тормозные – оказывают тормозное влияние на эфферентные нейроны.
В зависимости от вида медиатора в синапсе нейрона различают:
холинергические нейроны (медиатор – ацетилхолин)
адренергические нейроны (медиаторы – адреналин и норадреналин)
Среди множества известных функций нейрона: трофической, генераторной, проводящей возбуждение, наибольший интерес представляет способность нейронов синтезировать и секретировать биологическиактивные вещества. Одна нервная клетка может синтезировать только один вид медиатора.
- Генерация биоэлектрических потенциалов. В ходе эволюции у железистого эпителия, мышечной и нервной тканей появилось свойство возбудимости – способность реагировать на воздействие окружающей среды возбуждением. Внешним проявлением возбуждения является возникновение биоэлектрического потенциала.
Все ткани организма могут находиться в двух состояниях:
состоянии относительного физиологического покоя;
состоянии активности.
Наблюдается при раздражении ткани. Существует 2 вида активного состояния тканей: возбуждение и торможение. Возбуждение – это активный процесс, представляющий собой ответную реакцию ткани на раздражение и характеризующийся повышением функций ткани. Возбуждение характеризуется двумя группами признаков: неспецифическими и специфическими.
Неспецифические признаки возникают у всех возбудимых тканей вне зависимости от их строения:
изменение проницаемости клеточных мембран
изменение заряда клеточных мембран,
повышение потребления кислорода
повышение температуры
усиление обменных процессов
Специфические признаки различаются у различных тканей:
мышечная ткань – сокращение
железистая ткань – выделение секрета
нервная ткань – генерация нервного импульса.
Процесс возбуждения связан с наличием в мембране электрически (для ионов кальция и хлора) и химически (для ионов натрия и калия) управляемых каналов, которые могут открываться в ответ на соответствующее раздражение клетки.
Ионоселективные каналы. Для каждого из переносимых через мембрану вида ионов существуют самостоятельные транспортные системы – ионные каналы (натриевые, калиевые, кальциевые, каналы для хлора и т.д.). Ионный канал состоит из поры, воротного механизма, сенсора (индикатора) напряжения ионов в самой мембране и селективного фильтра.
Пора представляет собой молекулярное динамическое образование, которое может находиться в открытом и закрытом состоянии. Образована пора «транспортным» ферментом – белком с высокой каталитической активностью, который способен переносить ионы через мембрану со скоростью в 200 раз превышающей скорость простой диффузии.
Воротный механизм (ворота канала) расположен на внутренней стороне мембраны и представлен белковыми молекулами, способными к конформации (изменение пространственной конфигурации молекул). В тысячные доли секунды он открывает (активирует) и закрывает (инактивирует) канал и таким образом регулирует скорость передвижения ионов по нему и поступление их в цитоплазму.
Сенсор напряжения ионов в мембране представлен белковой молекулой, расположенной в самой мембране и способной реагировать на изменение мембранного потенциала.
Селективный фильтр находится в самом узком месте канала. Он определяет однонаправленное движение ионов через пору и ее избирательную проницаемость.
В развитии возбуждения выделяют 4 этапа:
1) предшествующее возбуждению состояние покоя (статическая поляризация);
2) деполяризацию;
3) реполяризацию
4) гиперполяризацию.
Статическая поляризация – наличие постоянной разности потенциалов между наружной и внутренней поверхностями клеточной мембраны. В состоянии покоя наружная поверхность клетки всегда электроположительна по отношению к внутренней, т.е. поляризована. Эта разность потенциалов, равная ~ 60 мВ, называется потенциалом покоя, или мембранным потенциалом (МП). В образовании потенциала принимают участие 4 вида ионов: катионы натрия (положительный заряд), катионы калия (положительный заряд), анионы хлора (отрицательный заряд), анионы органических соединений (отрицательный заряд). Во внеклеточной жидкости высока концентрация ионов натрия и хлора, во внутриклеточной жидкости – ионов калия и органических соединений. В состоянии относительного физиологического покоя клеточная мембрана хорошо проницаема для катионов калия, чуть хуже для анионов хлора, практически непроницаема для катионов натрия и совершенно непроницаема для анионов органических соединений.
В покое ионы калия без затрат энергии выходят в область меньшей концентрации (на наружную поверхность клеточной мембраны), неся с собой положительный заряд. Ионы хлора проникают внутрь клетки, неся отрицательный заряд. Ионы натрия продолжают оставаться на наружной поверхности мембраны, еще больше усиливая положительный заряд.
Деполяризация – сдвиг МП в сторону его уменьшения. Под действием раздражения открываются «быстрые» натриевые каналы, вследствие чего ионы Na лавинообразно поступают в клетку. Переход положительно заряженных ионов в клетку вызывает уменьшение положительного заряда на ее наружной поверхности и увеличение его в цитоплазме. В результате этого сокращается трансмембранная разность потенциалов, значение МП падает до 0, а затем по мере дальнейшего поступления Na в клетку происходят перезарядка мембраны и инверсия ее заряда (поверхность становится электроотрицательной по отношению к цитоплазме) – возникает потенциал действия (ПД). Электрографическим проявлением деполяризации является спайк, илипиковый потенциал.
Во время деполяризации, когда переносимый ионами Na положительный заряд достигает некоторого порогового значения, в сенсоре напряжения ионных каналов возникает ток смещения, который «захлопывает» ворота и «запирает» (инактивирует) канал, прекращая тем самым дальнейшее поступление Na в цитоплазму. Канал «закрыт» (инактивирован) вплоть до восстановления исходного уровня МП.
Реполяризация – восстановление исходного уровня МП. При этом ионы натрия перестают проникать в клетку, проницаемость мембраны для калия увеличивается, и он достаточно быстро выходит из нее. В результате заряд клеточной мембраны приближается к исходному. Электрографическим проявлением реполяризации является отрицательный следовой потенциал.
Гиперполяризация – увеличение уровня МП. Вслед за восстановлением исходного значения МП (реполяризация) происходит его кратковременное увеличение по сравнению с уровнем покоя, обусловленное повышением проницаемости калиевых каналов и каналов для Cl . В связи с этим поверхность мембраны приобретает избыточный по сравнению с нормой положительный заряд, а уровень МП становится несколько выше исходного. Электрографическим проявлением гиперполяризации является положительный следовой потенциал. На этом заканчивается одиночный цикл возбуждения.
Физиологические свойства нервных центров: пространственная и временная суммация возбуждений, трансформация ритма, посттетаническая потенциация, низкая лабильность, утомляемость, чувствительность к неиротропным средствам.
Нервным центром (НЦ) называется совокупность нейронов в различных отделах ЦНС, обеспечивающих регуляцию какой-либо функции организма. Например, бульбарный дыхательный центр.
Для проведения возбуждения через нервные центры характерны следующие особенности:
1. Одностороннее проведение. Оно идет от афферентного, через вставочный, к эфферентному нейрону. Это обусловлено наличием межнейронных синапсов.
2. Центральная задержка проведение возбуждения. Т.е. по НЦ возбуждение идет значительно медленнее, чем по нервному волокну. Это объясняется синаптической задержкой. Так как больше всего синапсов в центральном звене рефлекторной дуги, там скорость проведения наименьшая. Исходя из этого, время рефлекса, это время от начала воздействия раздражителя до появления ответной реакции. Чем длительнее центральная задержка, тем больше время рефлекса. Вместе с тем оно зависит от силы раздражителя. Чем она больше, тем время рефлекса короче и наоборот. Это объясняется явлением суммации возбуждений в синапсах. Кроме того, оно определяется и функциональным состоянием ЦНС. Например, при утомлении НЦ длительность рефлекторной реакции увеличивается.
3. Пространственная и временная суммация. Временная суммация возникает, как и в синапсах вследствие того, что чем больше поступает нервных импульсов, тем больше выделяется нейромедиатора в них, тем выше амплитуда ВПСП. Поэтому рефлекторная реакция может возникать на несколько последовательных подпороговых раздражений. Пространственная суммация наблюдается тогда, когда к нервному центру идут импульсы от нескольких рецепторов нейронов. При действии на них подпороговых стимулов, возникающие постсинаптические потенциалы суммируются и в мембране нейрона генерируется распространяющийся ПД.
4. Трансформация ритма возбуждения – изменение частоты нервных импульсов при прохождении через нервный центр. Частота может понижаться или повышаться. Например, повышающая трансформация (увеличение частоты) обусловлено дисперсией и мультипликацией возбуждения в нейронах. Первое явление возникает в результате разделения нервных импульсов на несколько нейронов, аксоны которых образуют затем синапсы на одном нейроне (рис). Второе, генерацией нескольких нервных импульсов при развитии возбуждающего постсинаптического потенциала на мембране одного нейрона. Понижающая трансформация объясняется суммацией нескольких ВПСП и возникновением одного ПД в нейроне.
5. Посттетаническая потенциация, это усиление рефлекторной реакции в результате длительного возбуждения нейронов центра. Под влиянием многих серий нервных импульсов, проходящих с большой частотой через синапсы,. выделяется большое количество нейромедиатора в межнейронных синапсах. Это приводит к прогрессирующему нарастанию амплитуды возбуждающего постсинаптического потенциала и длительному (несколько часов) возбуждению нейронов.
6. Последействие, это запаздывание окончания рефлекторного ответа после прекращения действия раздражителя. Связано с циркуляцией нервных импульсов по замкнутым цепям нейронов.
7. Тонус нервных центров – состояние постоянной повышенной активности. Он обусловлен постоянным поступлением к НЦ нервных импульсов от периферических рецепторов, возбуждающим влиянием на нейроны продуктов метаболизма и других гуморальных факторов. Например проявлением тонуса соответствующих центров является тонус определенной группы мышц.
8. Автоматия или спонтанная активность нервных центров. Периодическая или постоянная генерация нейронами нервных импульсов, которые возникают в них самопроизвольно, т.е. в отсутствии сигналов от других нейронов или рецепторов. Обусловлена колебаниями процессов метаболизма в нейронах и действием на них гуморальных факторов.
9. Пластичность нервных центров. Это их способность изменять функциональные свойства. При этом центр приобретает возможность выполнять новые функции или восстанавливать старые после повреждения. В основе пластичности Н.Ц. лежит пластичность синапсов и мембран нейронов, которые могут изменять свою молекулярную структуру.
10. Низкая физиологическая лабильность и быстрая утомляемость. Н.Ц. могут проводить импульсы лишь ограниченной частоты. Их утомление объясняется утомлением синапсов и ухудшением метаболизма нейронов.
18. Физиологические свойства мышц. Типы мышечных сокращений. Закон силы.
В организме имеются 3 типа мышц: скелетные или поперечно-полосатые, гладкие и сердечная. Скелетные мышцы обеспечивают перемещение тела в пространстве, поддержание позы тела за счет тонуса мышц конечностей и тела. Гладкие мышцы необходимы для перистальтики органов желудочно-кишечного тракта, мочевыводящей системы, регуляции тонуса сосудов, бронхов и т.д. Сердечная мышца служит для сокращения сердца и перекачивания крови. Все мышцы обладают возбудимостью, проводимостью и сократимостью, а сердечная и многие гладкие мышцы автоматией -способностью к самопроизвольным сокращениям.
При изучении различных типов сокращения пользуются тем. что мышца сохраняет способность сокращаться и после того, как она выде-пена из организма. Для экспериментальных целей обычно используют икроножную мышцу лягушки; если поддерживать эту мышцу во влажном состоянии, она может сокращаться часами. Для того чтобы регистрировать сокращения, один конец мышцы (ее начало) закрепляют неподвижно, а другой конец (с сухожилием) соединяют с подвижным рычажком, имеющим заостренный кончик . Этот кончик соприкасается с цилиндром, покрытым закопченной бумагой и приводимым во вращение часовым механизмом, так что мышца, приподнимая рычажок, регистрирует силу и продолжительность каждого своего сокращения. Можно применять дополнительные записывающие приспособления для отметки соответствующей шкалы времени и регистрации момента раздражения.
Одиночное сокращение. Когда мышца получает единичный стимул (например, единичный электрический импульс), она отвечает быстрым одиночным сокращением, продолжительность которого для мышцы лягушки составляет примерно 0,1 сек, а для мышцы человека — примерно 0,05 сек. Лабораторная запись одиночно-го сокращения показывает, что оно состоит из трех отдельных фаз: 1) латентного периода продолжительностью около 0,01 сек — промежутка времени между воздействием стимула и началом видимого укорочения мышцы; 2) периода сокращения продолжительностью около 0,04 сек, в течение которого мышца укорачивается и производит работу; 3) периода расслабления, самого продолжительного из трех (0,05 сек), во время которого мышца возвращается к своей первоначальной длине.
Скелетным мышечным волокнам, так же как и нервным волокнам (см. разд. 250), свойствен рефрактерный период — очень короткий, следующий непосредственно за раздражением период, во время которого они не способны отвечать на второе раздражение. Рефрактерный период скелетной мышцы настолько краток (около 0,002 сек), что мышца уже может реагировать на второе раздражение, когда она еще сокращается под действием первого. Наложение второго сокращения на первое приводит к более сильному укорочению мышечного волокна — явлению, называемому суммацией. Первичный эффект раздражения мышцы состоит в возникновении и распространении электрической реакции — мышечного потенциала действия, за которым следует изменение в структуре сократимого белка, актоми-озина, выявляемое по изменению двойного лучепреломления мышцы. После сокращения мышца потребляет больше кислорода и выделяет больше углекислоты и тепла, чем во время покоя; это так называемый период восстановления, в течение которого мышца возвращается- в исходное состояние. Период восстановления продолжается несколько секунд, и если мышцу повторно раздражать таким образом, чтобы последующие сокращения происходили раньше, чем мышца вернется в нормальное состояние после предшествующих, то мышца утомляется, сокращения становятся слабее и, наконец, прекращаются. Если утомленной мышце дать некоторое время отдохнуть, она вновь приобретает способность сокращаться.
Тетанус. Нормальное сокращение мышцы протекает не в форме одиночных сокращений, а в форме длительного сокращенного состояния, вызываемого «залпом» отдельных нервных импульсов, приходящих к мышце в быстрой последовательности. Такого рода длительное сокращение называется тетанусом1; во время тетануса нервные импульсы поступают с такой частотой (несколько сот в секунду), что расслабление между последовательными сокращениями произойти не успевает. В большинстве случаев при тетаническом сокращении отдельные мышечные волокна получают импульс не одновременно, а, так сказать, посменно; хотя отдельные волокна сокращаются и расслабляются, мышца как целое остается частично сокращенной. Каждый знает, что любая мышца тела может сократиться в различной степени. Эта градация сокращений регулируется нервной системой: при одновременном возбуждении лишь небольшой доли мышечных волокон получается слабое сокращение, а при одновременном возбуждении большого их числа — более сильное сокращение.
Тонус. Тонусом называется состояние длительного частичного сокращения, в котором находятся все нормальные скелетные мышцы, пока не повреждены идущие к ним нервы. Сер-
Тетанус не следует смешивать танией, т. е, мышечными спазмами, наступающими при недостатке гормона паращитовидных желез, и с болезнью тетанусом (латинское название столбняка), характеризующейся патологическим сокращением мышц и вызываемой И столбнячными бациллами. | z