

2. Первичная и вторичная структура днк.
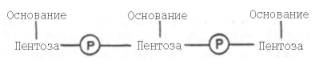
Под первичной структурой ДНК понимают порядок, последовательность расположения мононуклеотидов в полинуклеотидной цепи . Такая цепь стабилизируется 3',5'-фосфодиэфирными связями. Во всех нуклеиновых кислотах (точнее, в одноцепочечной нуклеиновой кислоте) имеется один и тот же тип связи – 3',5'-фосфодиэфирная связь между соседними нуклеотидами. Эту общую основу структуры можно представить следующим образом:
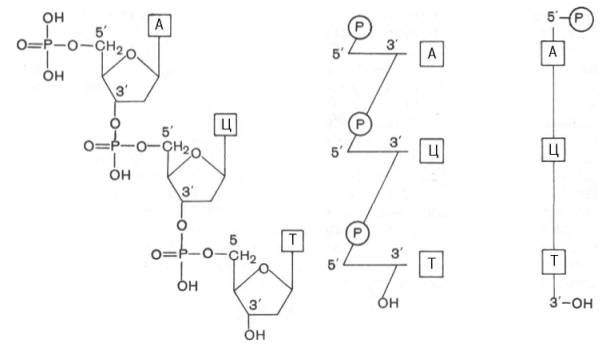
Установлено, что в образовании межнуклеотидной связи участвуют гидроксильные группы в 3'- и 5'-положениях остатков углевода.
В последнее время о первичной структуре ДНК (точнее, отдельных ее фрагментов) судят по ряду косвенных данных, например, по степени сплоченности нуклеотидных звеньев в молекуле ДНК (определение сводится в конечном счете к выяснению числа и структуры отдельных фракций нуклеотидов, так называемых изоплитов), также по кинетике реассоциации ДНК (метод позволяет выяснить наличие в молекуле повторяющихся последовательностейнуклеотидов). О первичной структуре ДНК судят, кроме того, по распределению минорных оснований (имеются данные о существовании подобной закономерности) и обнаружению в ДНК и определению последовательности палиндромов («обратно бегущие» последовательности, или перевертыши), которые обнаруживаются главным образом в местах рестрикции . Большие надежды в определении первичной структуры ДНК исследователи возлагают на физические, химические (синтез генов), генетические и другие методы, а также на методы выделения некоторых генов (или их фрагментов) из природных источников и синтеза генов на мРНК при участии ферментаобратной транскриптазы.
В соответствии с моделью Дж. Уотсона и Ф. Крика, предложенной в 1953 г. на основании ряда аналитических данных, а также рентгеноструктурного анализа молекула ДНК состоит из двух цепей, образуя правовращающую спираль, в которую обе полинуклеотидные цепи закручены вокруг одной и той же оси. Удерживаются цепи благодаря водородным связям, образующимся между их азотистыми основаниями .Обе цепи поли-нуклеотидов в биспиральной молекуле ДНК имеют строго определенное пространственное расположение, при котором азотистые основания находятся внутри, а фосфорильные и углеводные компоненты – снаружи.
Детальный анализ всевозможных вариантов образования водородных связей между основаниями показал, что в биспиральной молекуле ДНК основания уложены парами: пурин из одной цепи и пиримидин из другой в соответствии с правилами Чаргаффа. Поскольку ориентация оснований на плоскости не является, очевидно, произвольной, и основания в поли-нуклеотидах представлены в лактамной форме, наиболее вероятными были признаны пары аденин–тимин и гуанин–цитозин. Этот способ спаривания получил в дальнейшем экспериментальное подтверждение. Избирательность взаимодействия пар А–Т и Г–Ц принято выражать термином «комплементарность», а соответствующие азотистые основания называют комплементарными. Стабильность А–Т оснований обеспечивается двумя водородными связями, а пар Г–Ц – тремя, что в свою очередь определяется особенностями расположения функциональных групп азотистых оснований. Длина водородных связей между основаниями составляет около 0,3 нм. Таким образом, комплементарными оказываются не только отдельные основания, но и дезоксирибонуклеотидные цепи ДНК в целом, способствующие образованию весьма компактной структуры и стабилизации всей молекулы.