

Глава 6. Основные среды жизни
Распределение организмов по средам жизни
В процессе длительного исторического развития живой материи и формирования все более совершенных форм живых существ, организмы, осваивая новые места обитания, распределились на Земле соответственно ее минеральным оболочкам (гидросфера, литосфера, атмосфера) и приспособились к существованию в строго определенных условиях.
Первой средой жизни стала вода. Именно в ней возникла жизнь. По мере исторического развития многие организмы начали заселять наземно-воздушную среду. В результате появились наземные растения и животные, которые бурно эволюционировали, адаптируясь к новым условиям существования.
В процессе функционирования живого вещества на суше поверхностные слои литосферы постепенно преобразовывались в почву, в своеобразное, по выражению В. И. Вернадского, биокосное тело планеты. Почву стали заселять как водные, так и наземные организмы, создавая специфичный комплекс ее обитателей.
По всей вероятности, параллельно шло формирование паразитов и симбионтов, средой жизни которых в воде, на суше и в почве стали другие организмы – хозяева и сожители.
Таким образом, на современной Земле четко выделяются четыре среды жизни – водная, наземно-воздушная, почва и живые организмы,– существенно различающиеся своими условиями. Рассмотрим каждую из них.
Водная среда
Общая характеристика. Водная среда жизни, гидросфера, занимает до 71 % площади земного шара. По объему запасы воды на Земле исчисляются в пределах 1370 млн. куб. км, что составляет 1/800 часть объема земного шара. Основное количество воды, более 98 %, сосредоточено в морях и океанах, 1,24 % представлено льдами полярных областей; в пресных водах рек, озер и болот количество воды не превышает 0,45 %.
В водной среде обитает около 150000 видов животных (примерно 7 % от общего их количества на земном шаре) и 10 000 видов растений (8 %). Несмотря на то что представители абсолютного большинства групп растений и животных остались в водной среде (в своей «колыбели»), число их видов значительно меньше, чем наземных. Значит, эволюция на суше проходила значительно быстрее.
Наиболее разнообразен и богат растительный и животный мир морей и океанов экваториальных и тропических областей (особенно Тихого и Атлантического океанов). На юг и север от этих поясов качественный состав организмов постепенно обедняется. В районе Ост-Индского архипелага распространено около 40000 видов животных, а в море Лаптевых всего 400. При этом основная масса организмов Мирового океана сосредоточена в сравнительно небольшой по площади зоне морских побережий умеренного пояса и среди мангровых зарослей тропических стран. На обширных акваториях вдали от берега расположены пустынные области, практически лишенные жизни.
Удельный вес рек, озер и болот по сравнению с таковым морей и океанов в биосфере незначительный. Тем не менее они создают необходимый для огромного количества растений и животных, а также для человека запас пресной воды.
Водная среда оказывает сильное влияние на ее обитателей. В свою очередь живое вещество гидросферы воздействует на среду обитания, перерабатывает ее, вовлекая в круговорот веществ. Подсчитано, что вода морей и океанов, рек и озер разлагается и восстанавливается в биотическом круговороте за 2 млн. лет, т. е. вся она прошла через живое вещество планеты не одну тысячу раз *. Таким образом, современная гидросфера представляет собой продукт жизнедеятельности живого вещества не только современной, но и прошлых геологических эпох.
Характерной чертой водной среды является ее подвижность даже в стоячих водоемах, не говоря уже о проточных, быстро текущих реках и ручьях. В морях и океанах наблюдаются приливы и отливы, мощные течения, штормы; в озерах вода перемещается под действием ветра и температуры. Движение воды обеспечивает снабжение водных организмов кислородом и питательными веществами, приводит к выравниванию (снижению) температуры во всем водоеме.
Обитатели водоемов выработали соответствующие приспособления к подвижности среды. К примеру, в проточных водоемах встречаются прочно прикрепленные к подводным предметам так называемые «обрастающие» растения ·– зеленые водоросли (Cladophora) со шлейфом отростков, диатомовые (Diatomeae), водяные мхи (Fontinalis), образующие плотный покров даже на камнях в бурных речных перекатах.
Животные также адаптировались к подвижности водной среды. У рыб, обитающих в быстро текущих реках, тело в поперечном разрезе почти округлое (форель, гольян). Двигаются они обычно навстречу течению. Беспозвоночные проточных водоемов обычно держатся на дне, тело их сплющено в дорзо-вентральном направлении, многие имеют на брюшной стороне различные органы фиксации, позволяющие им прикрепляться к подводным предметам. В морях наиболее сильное влияние подвижных масс воды испытывают организмы приливно-отливной и прибойной зон. На скалистых берегах в прибойной полосе распространены прочно удерживающиеся на камнях усоногие раки (Balanus, Chthamalus), брюхоногие моллюски (Patella Haliotis), некоторые виды ракообразных, укрывающихся в расщелинах берега.
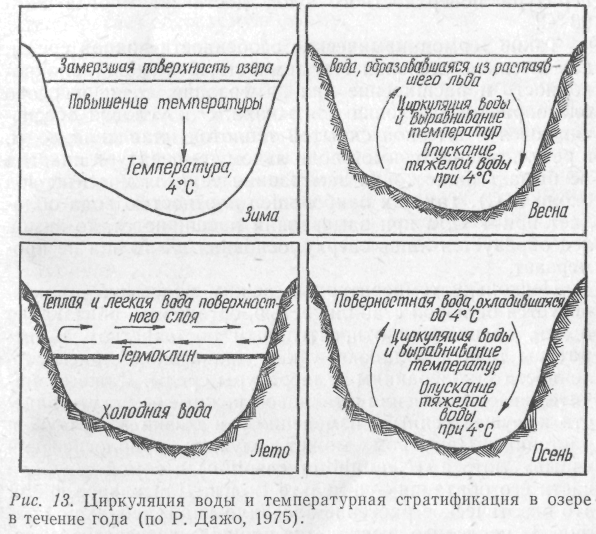
В жизни водных организмов умеренных широт большую роль играет вертикальное перемещение воды в стоячих водоемах. Вода в них четко делится на три слоя: верхний–эпилимнион, температура которого испытывает резкие сезонные колебания; слой температурного скачка– металимнион (термоклин), где наблюдается резкий перепад температур; придонный глубинный слой, гиполимнион,– здесь температура в течение года изменяется незначительно.
В летнее время наиболее теплые слои воды располагаются у поверхности, а холодные – у дна. Такое послойное распределение температур в водоеме называется прямой стратификацией. Зимой, с понижением температуры, наблюдается обратная стратификация: поверхностные холодные воды с температурой ниже 4 °С располагаются над сравнительно теплыми. Это явление называется температурной дихотомией. Оно особенно выражено в большинстве наших озер летом и зимой. В результате температурной дихотомии в водоеме образуется плотностная стратификация воды, нарушается вертикальная циркуляция ее и наступает период временного застоя –-стагнация.
Весной поверхностная вода вследствие нагревания до 4 °С становится более плотной и погружается вглубь, а на ее место с глубины поднимается более теплая вода. В результате такой вертикальной циркуляции в водоеме наступает гомотермия, т. е. на какое-то время температура всей водной массы выравнивается. С дальнейшим повышением температуры верхние слои воды становятся все менее плотными и уже не опускаются – наступает летняя стагнация.
Осенью же поверхностный слой охлаждается, становится более плотным и опускается вглубь, вытесняя на поверхность более теплую воду. Это происходит до наступления осенней гомотермии. При охлаждении поверхностных вод ниже 4 °С они снова становятся менее плотными и опять остаются на поверхности. В результате прекращается циркуляция воды и наступает зимняя стагнация.
Организмы в водоемах умеренных широт хорошо приспособлены к сезонным вертикальным перемещениям слоев воды, к весенней и осенней гомотермии и к летней и зимней стагнации (рис. 13).
В озерах тропических широт температура воды на поверхности никогда не опускается ниже 4 °С и температурный градиент в них четко выражен до самых глубинных слоев. Перемешивание воды, как правило, происходит здесь нерегулярно в наиболее холодное время года.
Своеобразные условия для жизни складываются не только в толще воды, но и на дне водоема, так как в грунтах отсутствует аэрация и из них вымываются минеральные соединения. Поэтому они не обладают плодородием и служат для водных организмов лишь более или менее твердым субстратом, выполняя в основном механико-динамическую функцию. В связи с этим наибольшее экологическое значение приобретают размеры частиц грунта, плотность их прилегания друг к другу и устойчивость к смыву течениями.
Абиотические факторы водной среды. Вода как среда жизни обладает особыми физико-химическими свойствами.
Температурный режим гидросферы коренным образом отличается от такового в других средах. Колебания температур в Мировом океане сравнительно невелики: самая низкая около –2 °С, а самая высокая приблизительно 36 °С. Амплитуда колебаний здесь, таким образом, укладывается в 38 °С. С глубиной температура воды в океанах падает. Даже в тропических областях на глубине 1000 м не превышает 4–5°С. На глубинах всех океанов залегает слой холодной воды (от -1,87 до +2°С).
В пресных внутренних водоемах умеренных широт температура поверхностных слоев воды колеблется от – 0,9 до +25°С, в более глубинных составляет 4–5 °С. Исключением являются термальные источники, где температура поверхностного слоя достигает иногда 85– 93 °С.
Такие термодинамические особенности водной среды, как высокая удельная теплоемкость, большая теплопроводность и расширение при замерзании, создают особо благоприятные условия для жизни. Эти условия обеспечиваются и высокой скрытой теплотой плавления воды, в результате чего зимой подо льдом температура никогда не бывает ниже точки замерзания ее (для пресных вод около 0°С). Так как наибольшей плотностью вода обладает при 4° С, а при замерзании расширяется, то зимой лед образуется лишь сверху, основная же толща не промерзает.
Поскольку температурный режим водоемов характеризуется большой стабильностью, организмы, обитающие в ней, отличаются относительным постоянством температуры тела и обладают узким диапазоном приспособленности к колебаниям температуры среды. Даже незначительные отклонения в тепловом режиме се могут привести к существенным изменениям в жизни животных и растений. Примером может служить «биологический взрыв» лотоса (Nelumbium caspium) в самой северной части его обитания – в дельте Волги. Длительное время это экзотическое растение заселяло лишь небольшой залив. За последнее десятилетие площадь лотосовых зарослей увеличилась почти в 20 раз и занимает теперь свыше 1500 га акватории. Такое бурное распространение лотоса объясняется общим падением уровня Каспийского моря, что сопровождалось образованием множества мелких озер и лиманов в устье Волги. В жаркие летние месяцы вода здесь прогревалась сильнее, чем прежде, это и способствовало разрастанию зарослей лотоса.
Воде свойственна также значительная плотность (в этом отношении она в 800 раз превосходит воздушную среду) и вязкость. На растениях эти особенности сказываются в том, что у них очень слабо или вовсе не развивается механическая ткань, поэтому стебли их очень эластичны и легко изгибаются. Большинству водных растений присуща плавучесть и способность находиться в толще воды во взвешенном состоянии. Они то поднимаются к поверхности, то вновь опускаются. У многих водных животных покровы обильно смазываются слизью, уменьшающей трение при передвижении, а тело приобретает обтекаемую форму.
Организмы в водной среде распределены по всей ее толще (в океанических впадинах животные обнаружены на глубинах свыше 10000 м). Естественно, что на разных глубинах они испытывают различное давление. Глубоководные приспособлены к высокому давлению (до 1000 атм), обитатели же поверхностных слоев ему не подвержены. В среднем в водной толще на каждые 10 м глубины давление возрастает на 1 атм. Все гидробионты приспособлены к этому фактору и соответственно делятся на глубоководных и обитающих на незначительных глубинах.
Большое влияние на водные организмы оказывают прозрачность воды и ее световой режим. Особенно это сказывается на распространении фотосинтезирующих растений. В мутных водоемах они обитают только в поверхностном слое, а там, где большая прозрачность, проникают на значительные глубины. Определенную мутность воды создает огромное количество взвешенных в ней частиц, что лимитирует проникновение солнечных лучей. Мутность воды может быть обусловлена частицами минеральных веществ (глина, ил), мелкими организмами. Уменьшается прозрачность воды и летом при бурном разрастании водной растительности, при массовом размножении мелких организмов, находящихся во взвешенном состоянии в поверхностных слоях. Световой режим водоемов зависит и от сезона. На севере в умеренных широтах, когда водоемы замерзают, а лед сверху еще покрывается снегом, проникновение света в толщу воды сильно ограничивается.
Световой режим обусловливается также закономерным убыванием света с глубиной из-за того, что вода поглощает солнечный свет. При этом лучи с разной длиной волны поглощаются неодинаково: быстрее всего–красные, тогда как сине-зеленые проникают на значительные глубины. В океане с глубиной становится все темнее. Цвет среды при этом меняется, постепенно переходя от зеленоватого до зеленого, затем до голубого, синего, сине-фиолетового, сменяемого постоянным мраком. Соответственно этому с глубиной зеленые водоросли (Chlorophyta) сменяются бурыми (Phaeophyta) и красными (Rhodophyta), пигменты которых приспособлены к улавливанию солнечных лучей с разной длиной волны. С глубиной также закономерно меняется и окраска животных. В поверхностных, светлых слоях воды обычно обитают ярко и разнообразно окрашенные животные, тогда как глубоководные виды лишены пигментов. В сумеречной зоне океана обитают животные, окрашенные в цвета с красноватым оттенком, что помогает им скрываться от врагов, так как красный цвет в сине-фиолетовых лучах воспринимается как черный.
Немаловажную роль в жизни водных организмов играет соленость воды. Как известно, вода является прекрасным растворителем многих минеральных соединений. В результате природным водоемам свойствен определенный химический состав. Наибольшее значение имеют карбонаты, сульфаты, хлориды. Количество растворенных солей на 1 л воды в пресных водоемах не превышает 0,5 г (обычно меньше), в морях и океанах оно достигает 35 г (табл. 6).
Таблица 6. Распределение основных солей в различных водоемах (по Р. Дажо, 1975)
Водоемы |
Сульфаты, % |
Хлориды, % |
Карбонаты, % |
Соленость, г/л |
Пресные воды |
13,2 |
6,9 |
79,9 |
_ |
Открытый океан |
10,8 |
88,8 |
0,4 |
35 |
Черное море |
9,69 |
80,71 |
2,59 |
19 |
Каспийское море |
30,5 |
63,36 |
1,24 |
12,86 |
Аральское море |
38,71 |
58,59 |
0,93 |
11,28 |
Для жизни пресноводных животных существенную роль играет кальций. Моллюски, ракообразные и другие беспозвоночные используют его для построения раковин, наружного скелета. Но пресные водоемы в зависимости от целого ряда обстоятельств (наличие тех или иных растворимых солей в грунте водоема, в грунте и почве берегов, в воде впадающих рек и ручьев) очень различаются как по составу, так и по концентрации растворенных в них солей. Морские воды более стабильны в этом отношении. В них обнаружены почти все известные элементы. Однако по степени значимости первое место занимает поваренная соль, затем хлористый и сернокислый магний и хлористый калий.
Пресноводные растения и животные обитают в гипотонической среде, т. е. в среде, в которой концентрация растворенных веществ ниже, чем в жидкостях тела и тканей. Из-за разницы в осмотическом давлении вне и внутри тела в организм постоянно проникает вода, и гидробионты пресных вод вынуждены интенсивно удалять ее. В связи с этим у них хорошо выражены процессы осморегуляции. Концентрация солей в жидкостях тела и тканей многих морских организмов изотонична концентрации растворенных солей в окружающей воде. Поэтому осморегуляторные функции у них не развиты в такой степени, как у пресноводных. Трудности осморегуляции являются одной из причин того, что многие морские растения и особенно животные не сумели заселить пресные водоемы и оказались, за исключением отдельных представителей, типичными морскими жителями (кишечнополостные – Coelenterata, иглокожие–·Echinodermata, погонофоры – Pogonophora, губки – Spongia, оболочники – Tunicata). В то же время в морях и океанах практически не обитают насекомые, тогда как пресноводные бассейны обильно ими заселены. Типично морские и типично пресноводные виды не переносят значительных изменений солености воды. Все они – стеногалинные организмы. Эвригалинных животных пресноводного и морского происхождения сравнительно немного. Обычно они встречаются, притом в значительных количествах, в солоноватых водах. Это пресноводный судак (Stizostedion lucioperca), лещ (Abramis brama), щука (Esox lucius), из морских можно назвать семейство кефалевых (Mugilidae).
В пресных водах распространены растения, укрепленные на дне водоема. Нередко их фотосинтетическая поверхность располагается над водой. Это рогозы (Typha), камыши (Scirpus), стрелолист (Sagittaria), кувшинки (Nymphaea), кубышки (Nuphar). У других фотосинтезирующие органы погружены в воду. К ним относятся рдесты (Potamogeton), уруть (Myriophyllum), элодея (Elodea). Некоторые высшие растения пресных вод лишены корней. Они либо свободно плавают, либо обрастают подводные предметы или водоросли, прикрепленные к грунту.
Если кислород для воздушной среды не играет существенной роли, то для водной – это важнейший экологический фактор. Содержание его в воде обратно пропорционально температуре. С понижением температуры растворимость кислорода, как и других газов, увеличивается. Накопление растворенного в воде кислорода происходит в результате поступления его из атмосферы, а также вследствие фотосинтетической деятельности зеленых растений. При перемешивании воды, что характерно для проточных водоемов и особенно для быстро текущих рек и ручьев, содержание кислорода также возрастает.
Различные животные проявляют неодинаковую потребность к кислороду. Например, форель (Salmo trutta), гольян (Phoxinus phoxinus) очень чувствительны к его дефициту и поэтому обитают лишь в быстро текущих холодных и хорошо перемешиваемых водах. Плотва (Rutilus rutilus), ерш (Acerina cernua), сазан (Cyprinus carpio), карась (Carassius carassius) неприхотливы в этом отношении, а личинки комаров хирономид (Chirono-midae) и малощетинковые черви трубочники (Tubifex) обитают на больших глубинах, где кислород вообще отсутствует или его очень мало. Водные насекомые и легочные моллюски (Pulmonata) также могут жить в водоемах с низким содержанием кислорода. Однако они систематически поднимаются к поверхности, запасая на некоторое время свежий воздух.
Углекислый газ растворяется в воде примерно в 35 раз лучше кислорода. В воде его почти в 700 раз больше, чем в атмосфере, откуда он поступает. Источником углекислоты в воде, кроме того, являются карбонаты и бикарбонаты щелочных и щелочноземельных металлов. Углекислый газ, содержащийся в воде, обеспечивает фотосинтез водных растений и принимает участие в формировании известковых скелетных образований беспозвоночных животных.
Большое значение в жизни водных организмов имеет концентрация водородных ионов (рН). Пресноводные бассейны с рН 3,7–4,7 считаются кислыми, 6,95–7,3 – нейтральными, с рН больше 7,8 – щелочными. В пресных водоемах рН испытывает даже суточные колебания. Морская вода более щелочная и рН ее значительно меньше изменяется, чем пресной. С глубиной рН уменьшается.
Концентрация водородных ионов играет большую роль в распределении гидробионтов. При рН меньше 7,5 растет полушник (Isoetes), ежеголовник (Sparganium), при 7,7–8,8, т. е. в щелочной среде, развиваются многие виды рдестов, элодея. В кислых водах болот преобладают сфагновые мхи (Sphagnum), но отсутствуют пластинчатожаберные моллюски из рода беззубок (Unio), редко встречаются другие моллюски, но в обилии распространены раковинные корненожки (Testacea). Большинство пресноводных рыб выдерживают рН от 5 до 9. Если рН меньше 5, наблюдается массовая гибель рыб, а выше 10 – погибают все рыбы и другие животные.
Экологические группы гидробионтов. Толща воды - пелагиаль (pelagos – море) заселена пелагическими организмами, способными активно плавать или удерживаться (парить) в определенных слоях. В соответствии с этим пелагические организмы подразделяются на две группы – нектон и планктон. Обитатели дна образуют третью экологическую группу организмов – бентос.
Нектон (nekios –· плавающий) – это совокупность пелагических активно передвигающихся животных, не имеющих непосредственной связи с дном. В основном это крупные животные, способные преодолевать большие расстояния и сильные водные течение. Для них характерна обтекаемая форма тела и хорошо развитые органы движения. Типичными нектонными организмами являются рыбы, кальмары, ластоногие, киты. В пресных водах, кроме рыб, к нектону относятся земноводные и активно перемещающиеся насекомые. Многие морские рыбы могут передвигаться в толще воды с огромной скоростью. Очень быстро, до 45–50 км/ч, плавают некоторые кальмары (Oegopsida), парусники (Istiopharidae) развивают скорость до 100–ПО км/ч, а меч-рыба (Xiphias glabius) – до 130 км/ч.
Планктон (planktos – парящий, блуждающий) – это совокупность пелагических организмов, не обладающих способностью к быстрым активным передвижениям. Планктонные организмы не могут противостоять течениям. В основном это мелкие животные – зоопланктон и растения – фитопланктон. В состав планктона периодически включаются и парящие в толще воды личинки многих животных.
Планктонные организмы располагаются либо на поверхности воды, либо на глубине или даже в придонном слое. Первые составляют особую группу – нейстон. Организмы же, часть тела которых находится в воде, а часть – над ее поверхностью, получили название плейстон. Таковы сифонофоры (Siphonophora), ряска (Lemna) и др.
Фитопланктон имеет большое значение в жизни водоемов, поскольку является основным продуцентом органического вещества. К нему относятся прежде всего диатомовые (Diatomeae) и зеленые (Chlorophyta) водоросли, растительные жгутиконосцы (Phytomastigina), перидинеи (Peridineae) и кокколитофориды (Coccolitophoridae). В северных водах Мирового океана преобладают диатомовые водоросли, а в тропических и субтропических – панцирные жгутиконосцы. В пресных водах, кроме диатомовых, распространены зеленые и сине-зеленые (Суаnophyta) водоросли.
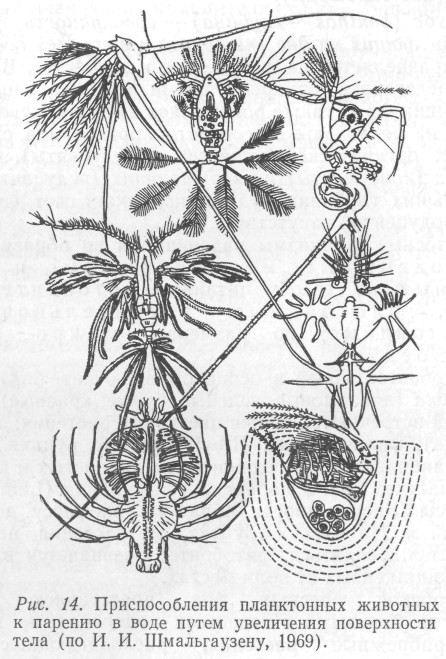
Зоопланктон и бактерии встречаются на всех глубинах. В морском зоопланктоне доминируют мелкие ракообразные (Copepoda, Amphipoda, Euphausiacea), простейшие (Foraminifera, Radiolaria, Tintinnoidea). Более крупные представители его – это крылоногие моллюски (Pteropoda), медузы (Scyphozoa) и плавающие гребневики (Ctenophora), сальпы (Salpae), некоторые черви (Alciopidae, Tomopteridae). В пресных водах распространены плохо плавающие сравнительно крупные ракообразные (Daphnia, Cyclopoidea, Ostracoda, Simocephalus; рис. 14), много коловраток (Rotatoria) и простейших.
Наибольшего видового разнообразия достигает планктон тропических вод.
Группы планктонных организмов различают по размеру. Наннопланктон (nannos – карликовый) наиболее мелкие водоросли и бактерии; микропланктон (micros – малый) – большинство водорослей, простейшие, коловратки; мезопланктон (mеsos – средний) – веслоногие и ветвистоусые раки, креветки и ряд животных и растений, не больше 1 см в длину; макропланктон (macros – крупный) –·медузы, мизиды, креветки и другие организмы, размером более 1 см; мегалопланктон (megalos– громадный) – очень крупные, свыше 1 м, животные. Например, плавающий гребневик венерин пояс (Cestus veneris) достигает в длину 1,5 м, а медуза цианея (Суапеа) имеет колокол диаметром до 2 м и щупальца длиною в 30 м.
Планктонные организмы являются важным пищевым компонентом многих водных животных (в том числе таких гигантов, как усатые киты – Mystacoceti), особенно если учесть, что для них, и прежде всего для фитопланктона, характерны сезонные вспышки массового размножения (цветение воды).
Бентос (benthos – глубина) – совокупность организмов, обитающих на дне (на грунте и в грунте) водоемов. Он подразделяется на фитобентос и зообентос. В основном представлен прикрепленными или медленно передвигающимися, а также роющимися в грунте животными. Только на мелководье он состоит из организмов, синтезирующих органическое вещество (продуценты), потребляющих (консументы) и разрушающих (редуценты) его. На больших глубинах, куда не проникает свет, фитобентос (продуценты) отсутствует.
Бентосные организмы различаются по образу жизни – подвижные, малоподвижные и неподвижные; по способу питания – фотосинтезирующие, плотоядные, растительноядные, детритоядные; по размерам – макро-, мезо- микробентос.
Фитобентос морей в основном включает бактерии и водоросли (диатомовые, зеленые, бурые, красные). У побережий встречаются также цветковые растения: зостера (Zostera), филлосподикс (Phyllospadix), руппия (Rup-pia). Наиболее богат фитобентос на скалистых и каменистых участках дна. У побережий ламинарии (Laminaria) и фукусы (Fucus) иногда образуют биомассу до 30 кг на 1 кв. м. На мягких грунтах, где растения не могут прочно прикрепляться, фитобентос развивается в основном в защищенных от волн местах.
Фитобенос пресных вод представлен бактериями, диатомовыми и зелеными водорослями. Обильны прибрежные растения, располагающиеся от берега вглубь четко выраженными поясами. В первом поясе произрастают полупогруженные растения (тростники, камыши, рогоз и осоки). Второй пояс занимают погруженные растения с плавающими листьями (кубышки, кувшинки, ряски, водокрас). В третьем поясе преобладают погруженные растения – рдесты, элодея и др.
Все водные растения по образу жизни можно разделить на две основные экологические группы: гидрофиты – растения, погруженные в воду только нижней частью и обычно укореняющиеся в грунте, и гидатофиты – растения, полностью погруженные в воду, но иногда плавающие на поверхности или имеющие плавающие листья.
В морском зообентосе доминируют фораминиферы, губки, кишечнополостные, немертины, многощетинковые черви, сипункулиды, мшанки, плеченогие, моллюски, асцидии, рыбы. Наиболее многочисленны бентосные формы на мелководьях, где общая биомасса их нередко достигает десятков килограммов на 1 кв. м. С глубиной численность бентоса резко падает и на больших глубинах составляет миллиграммы на 1 кв. м.
В. пресных водоемах зообентоса меньше, чем в морях и океанах, а видовой состав однообразнее. В основном это простейшие, некоторые губки, ресничные и малощетинковые черви, пиявки, мшанки, моллюски и личинки насекомых.
Экологическая пластичность водных организмов. Водные организмы обладают меньшей экологической пластичностью, чем наземные, так как вода – более стабильная среда и абиотические факторы ее претерпевают сравнительно незначительные колебания. Наименее пластичны морские растения и животные. Они очень чувствительны к изменениям солености воды и ее температуры. Так, мадрепоровые кораллы не выдерживают даже слабого опреснения воды и обитают только в морях, притом на твердом грунте при температуре не ниже 20 °С. Это типичные стенобионты. Однако есть виды и с повышенной экологической пластичностью. Например, корненожка Cyphoderia ampulla является типичным эврибионтом. Она обитает в морях и в пресных водах, в теплых прудах и в холодных озерах.
Пресноводные животные и растения, как правило, гораздо пластичнее, чем морские, поскольку пресная вода как среда жизни более изменчива. Самыми же пластичными являются солоноватоводные обитатели. Они адаптированы и к высокой концентрации растворенных солей и к значительному опреснению. Однако их насчитывается сравнительно небольшое количество видов, поскольку в солоноватых водах экологические факторы претерпевают существенные изменения.
Широту экологической пластичности гидробионтов оценивают по отношению не только ко всему комплексу факторов (эври- и станобионтность), но и к какому-либо одному из них. Прибрежные растения и животные в отличие от обитателей открытых зон в основном эвритермные и эвригалинные организмы, так как вблизи берега температурные условия и солевой режим довольно изменчивы (прогреваемость солнцем и сравнительно интенсивное охлаждение, опреснение притоком воды из ручьев и рек, особенно в период дождей, и т. д.). Типичным стенотермным видом является лотос. Он произрастает только в хорошо прогреваемых мелких водоемах. По этим же причинам более эвритермными и эвригалинными оказываются обитатели поверхностных слоев в сравнении с глубоководными формами.
Экологическая пластичность служит важным регулятором расселения организмов. Как правило, гидробионты с высокой экологической пластичностью распространены довольно широко. Это касается, к примеру, элодеи. Однако диаметрально противоположен ей в этом смысле рачок артемия (Artemia salina). Он живет в небольших водоемах с очень соленой водой. Это типичный стеногалинный представитель с узкой экологической пластичностью. Но по отношению к другим факторам он весьма пластичен и поэтому в соленых водоемах встречается повсеместно.
Экологическая пластичность зависит от возраста и фазы развития организма. Так, морской брюхоногий моллюск Littorina во взрослом состоянии ежедневно при отливах длительное время обходится без воды, а его личинки ведут сугубо планктонный образ жизни и не переносят высыхания.
Адаптивные особенности водных растений. Экология водных растений, как было отмечено, весьма специфична и резко отличается от экологии большинства наземных растительных организмов. Способность водных растений поглощать влагу и минеральные соли непосредственно из окружающей среды отражается на их морфологической и физиологической организации. Для водных растений прежде всего характерно слабое развитие проводящей ткани и корневой системы. Последняя служит в основном для прикрепления к подводному субстрату и в отличие от наземных растений не выполняет функцию минерального питания и водоснабжения. В связи с этим корни укореняющихся водных растений лишены корневых волосков. Питание их осуществляется всей поверхностью тела. Мощно развитые корневища у некоторых из них служат для вегетативного размножения и запасания питательных веществ. Таковы многие рдесты, кувшинки, кубышки.
Большая плотность воды обусловливает возможность обитания растений во всей ее толще. Для этого у низших растений, заселяющих различные слои и ведущих плавающий образ жизни, имеются специальные придатки, увеличивающие их плавучесть и позволяющие им удерживаться во взвешенном состоянии. У высших гидрофитов слабо развивается механическая ткань. В их листьях, стеблях, корнях, как отмечалось, располагаются воздухоносные межклеточные полости. Это увеличивает легкость и плавучесть взвешенных в воде и плавающих на поверхности органов, а также способствует смыванию внутренних клеток водой с растворенными в ней газами и солями. Гидатофиты в целом характеризуются большой поверхностью листьев при незначительном общем объеме растения. Это обеспечивает им интенсивный газообмен при недостатке растворенного в воде кислорода и других газов. У многих рдестов (Potamogeton lusens, P. perfoliatus) стебли и листья тонкие и очень длинные, покровы их легко проницаемы для кислорода. Другие растения обладают сильно расчлененными листьями (водяной лютик– Ranunculus aquatilis, уруть – Myriophyllum spicatum, роголистник – Ceratophyllum dernersum).
У ряда водных растений развита гетерофилия (разнолистность). Например, у сальвинии (Salvinia) погруженные листья выполняют функцию минерального питания, а плавающие – органического. У кувшинки и кубышки плавающие и погруженные в воду листья существенно отличаются друг от друга. Верхняя поверхность плавающих листьев плотная и кожистая с большим количеством устьиц. Это способствует лучшему газообмену с воздухом. На нижней стороне плавающих и на подводных листьях устьиц совсем нет.
Не менее важной адаптивной особенностью растений к обитанию в водной среде является и то, что погруженные в воду листья обычно очень тонкие. Хлорофилл в них часто располагается в клетках эпидермиса. Это приводит к усилению интенсивности фотосинтеза в условиях слабого освещения. Наиболее четко такие анатомо-морфологические особенности выражены у многих рдестов (Potamogeton), элодеи (Helodea canadensis), водяных мхов (Riccia, Fontinalis), валиснерии (Vallisneria spiralis).
Защитой водных растений от вымывания из клеток минеральных солей (выщелачивания) является выделение специальными клетками слизи и образование эндодермы в виде кольца из более толстостенных клеток.
Сравнительно низкая температура водной среды обусловливает отмирание вегетирующих частей у погруженных в воду растений после образования зимних почек, а также замену летних нежных тонких листьев более жесткими и короткими зимними. Вместе с тем низкая температура воды отрицательно влияет на генеративные органы водных растений, а высокая ее плотность затрудняет перенос пыльцы. Поэтому водные растения интенсивно размножаются вегетативным путем. Половой процесс у многих из них подавлен. Адаптируясь к особенностям водной среды, большинство погруженных и плавающих на поверхности растений выносят цветоносные стебли в воздушную среду и размножаются половым путем (пыльца разносится ветром и поверхностными течениями). Образующиеся плоды, семена и другие зачатки также распространяются поверхностными течениями (гидрохория).
К гидрохорным принадлежат не только водные, но и многие прибрежные растения. Плоды их обладают высокой плавучестью и могут длительное время находиться в воде, не теряя всхожести. Водой переносятся плоды и семена частухи (Alisma plantago-aquatica), стрелолиста (Sagittaria sagittifolia), сусака (Butomus umbellatus), рдестов и других растений. Плоды многих осок (Сагех) заключены в своеобразные мешочки с воздухом и тоже разносятся водными течениями. Предполагают, что даже кокосовые пальмы расселились по архипелагам тропических островов Тихого океана благодаря плавучести своих плодов – кокосовых орехов. Вдоль реки Вахш по каналам таким же образом расселился сорняк гумай (Sorgnum halepense).
Адаптивные особенности водных животных. Адаптации животных к водной среде еще более многообразны, чем растений. У них можно выделить анатомо-морфологические, физиологические, поведенческие и другие адаптивные признаки. Даже простое перечисление их затруднительно. Поэтому назовем в общих чертах лишь наиболее характерные из них.
Животные, обитающие в толще воды, обладают прежде всего приспособлениями, увеличивающими их плавучесть и позволяющими им противостоять движению воды, течениям. Донные организмы, наоборот, вырабатывают приспособления, препятствующие поднятию их в толщу воды, т. е. уменьшающие плавучесть и позволяющие удерживаться на дне даже в быстро текущих водах.
У мелких форм, живущих в толще воды, наблюдается редукция скелетных образований. У простейших (Rhizopoda, Radiolaria) раковины обладают пористостью, кремневые иглы скелета полые внутри. Удельная плотность медуз (Scyphozoa) и гребневиков (Ctenophora) уменьшается за счет наличия воды в тканях. Увеличение плавучести достигается и скоплением капелек жира в теле (ночесветки – Noctiluca, радиолярии – Radiolaria). Более крупные скопления жира наблюдаются и у некоторых ракообразных (Cladocera, Copepoda), рыб, китообразных. Удельную плотность тела снижают и газовые пузырьки в протоплазме раковинных амеб, воздухоносные камеры в раковинах моллюсков. Плавательные пузыри, наполненные газом, имеют многие рыбы. У сифонофор Physalia и Velella развиваются мощные воздухоносные полости.
Для пассивно плавающих в толще воды животных характерно не только уменьшение веса, но и увеличение удельной поверхности тела. Дело в том, что чем больше вязкость среды и выше удельная поверхность тела организма, тем медленнее он погружается в воду. В результате у животных уплощается тело, на нем образуются всевозможные шипы, выросты, придатки. Это свойственно многим радиоляриям (Chalengeridae, Aulacantha), жгутиковым (Leptodiscus, Craspedotella), фораминиферам (Globigerina, Orbulina). Поскольку с повышением температуры вязкость воды уменьшается, а с повышением солености – увеличивается, адаптации к увеличению трения наиболее резко выражены при высоких температурах и низких соленостях. Например, жгутиковые Ceratium из Индийского океана вооружены более длинными роговидными придатками, чем те, которые обитают в холодных водах Восточной Атлантики.
Активное плавание у животных осуществляется с помощью ресничек, жгутиков, изгибания тела. Так передвигаются простейшие, ресничные черви, коловратки.
Среди водных животных распространено плавание реактивным способом за счет энергии выбрасываемой струи воды. Это характерно для простейших, медуз, личинок стрекоз, некоторых двустворчатых моллюсков. Наивысшего совершенства реактивный способ передвижения достигает у головоногих моллюсков. Некоторые кальмары при выбрасывании воды развивают скорость 40–50 км/ч. У более крупных животных образуются специализированные конечности (плавательные ноги у насекомых, ракообразных; плавники, ласты). Тело у таких животных покрыто слизью и имеет обтекаемую форму.
Большая группа животных, в основном пресноводных, при передвижении использует поверхностную пленку воды (поверхностное натяжение). По ней свободно бегают, например, жуки вертячки (Gyrinidae), клопы водомерки (Gerridae, Veliidae). По нижней поверхности пленки передвигаются мелкие жуки Hydrophilidae, к ней подвешиваются и моллюски прудовики (Limnaea), личинки комаров. Все они обладают рядом особенностей в строении конечностей, а покровы их не смачиваются водой.
Только в водной среде встречаются неподвижные, ведущие прикрепленный образ жизни животные. Им свойственны своеобразная форма тела, незначительная плавучесть (плотность тела больше плотности воды) и специальные приспособления для прикрепления к субстрату. Одни прикрепляются к грунту, другие ползают по нему или ведут роющий образ жизни, некоторые поселяются на подводных предметах, в частности днищах кораблей.
Из прикрепленных к грунту животных наиболее характерны губки, многие кишечнополостные, особенно гидроиды (Hydroidea) и коралловые полипы (Anthozoa), морские лилии (Crinoidea), двустворчатые моллюски (Bivalvia), усоногие раки (Cirripedia) и др.
Среди роющих животных особенно много червей, личинок насекомых, а также моллюсков. Значительное время в грунте проводят определенные рыбы (шиповка – Cobitis taenia, камбаловые – Pleuronectidae, скаты – Rajidae), личинки миног (Petromyzones). Обилие этих животных и их видовое разнообразие зависят от типа грунта (камни, песок, глина, ил). На каменистых грунтах их обычно меньше, чем на илистых. Беспозвоночные, в массе заселяющие илистые грунты, создают оптимальные условия для жизни ряда более крупных придонных хищников.
Большинство водных животных пойкилотермны, и температура их тела зависит от температуры окружающей среды. У гомойотермных же млекопитающих (ластоногие, китообразные) образуется мощный слой подкожного жира, выполняющий теплоизоляционную функцию.
Для водных животных имеет значение давление среды обитания. В связи с этим выделяются стенобатные животные, не выдерживающие больших колебаний давления, и эврибатные, обитающие и при высоком и при низком давлении. Голотурии (Elpidia, Myriotrochus) живут на глубине от 100 до 9000 м, а многие виды раков Storthyngura, погонофоры, морские лилии располагаются на глубинах от 3000 до 10 000 м. У таких глубоководных животных отмечаются специфичные черты организации: увеличение размеров тела; исчезновение или слабое развитие известкового скелета; часто – редукция органов зрения; усиление развития осязательных рецепторов; отсутствие пигментации тела или, наоборот, темная окраска.
Поддержание определенного осмотического давления и ионного состояния растворов в теле животных обеспечивается сложными механизмами водно-солевого обмена. Однако большинство водных организмов пойкилосмотичны, т. е. осмотическое давление в их теле зависит от концентрации растворенных солей в окружающей воде. Гомойосмотичны лишь позвоночные, высшие раки, насекомые и их личинки – они сохраняют постоянное осмотическое давление в теле, независимо от солености воды.
Морские беспозвоночные в основном не имеют механизмов водно-солевого обмена: анатомически они закрыты для воды, а осмотически открыты. Однако говорить об абсолютном отсутствии у них механизмов, управляющих водно-солевым обменом, было бы неверно.
Они просто несовершенны, и это объясняется тем, что соленость морской воды близка к солености соков тела. Ведь у гидробионтов пресных вод соленость и ионное состояние минеральных веществ соков тела, как правило, выше, чем окружающей воды. Поэтому у них хорошо выражены механизмы осморегуляцни. Наиболее распространенным способом поддержания постоянного осмотического давления является регулярное удаление поступающей в организм воды с помощью пульсирующих вакуолей и органов выделения. У других животных для этих целей развиваются непроницаемые покровы из хитина или роговых образований. У некоторых на поверхности тела выделяется слизь.
Трудностью регулирования осмотического давления у пресноводных организмов объясняется их видовая бедность по сравнению с обитателями моря.
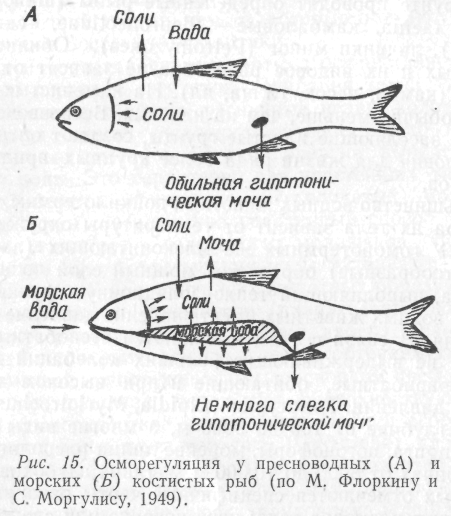
Проследим на примере рыб, как осуществляется осморегуляция животных в морских и пресных водах. Пресноводные рыбы избыток воды удаляют усиленной работой выделительной системы, а соли поглощают через жаберные лепестки. Морские рыбы, наоборот, вынуждены пополнять запасы воды и поэтому пьют морскую воду, а излишки поступающих с ней солей выводят из организма через жаберные лепестки (рис. 15).
Смена условий в водной среде вызывает определенные поведенческие реакции организмов. С изменением освещенности, температуры, солености, газового режима и других факторов связаны вертикальные миграции животных. В морях и океанах в таких миграциях (опускание вглубь, поднятие к поверхности) принимают участие миллионы тонн гидробионтов. При горизонтальных миграциях водные животные могут преодолевать сотни и тысячи километров. Таковы нерестовые, зимовальные и нагульные миграции многих рыб, водных млекопитающих.
Биофильтраторы и их экологическая роль. Одной из специфических особенностей водной среды является наличие в ней большого количества мелких частиц органического вещества – детрита, образующегося за счет отмирающих растений и животных. Огромные массы этих частиц оседают на бактериях и благодаря выделяющемуся в результате бактериального процесса газу постоянно находятся в толще воды во взвешенном состоянии.
Детрит для многих водных организмов представляет собой высококачественную пищу, поэтому некоторые из них, так называемые биофильтраторы, приспособились добывать его с помощью специфических микропористых структур. Эти структуры как бы отцеживают воду, задерживая взвешенные в ней частицы. Такой способ питания называется фильтрацией. Другая группа животных осаждает детрит на поверхность либо собственного тела, либо на специальные ловчие устройства. Такой способ называется седиментацией. Часто один и тот же организм питается путем и фильтрации и седиментации.
Животные-биофильтраторы (пластинчатожаберные моллюски, сидячие иглокожие и многощетинковые кольчецы, мшанки, асцидии, планктонные ракообразные и многие другие) играют большую роль в биологической очистке водоемов. Например, колония мидий (Mytilus) на 1 кв. м пропускает через мантийную полость до 250 куб. м воды в сутки, профильтровывая ее и осаждая взвешенные частицы. Почти микроскопический рачок каланус (Calanoida) очищает в сутки до 1,5 л воды. Если учесть громадную численность этих ракообразных, то работа, выполняемая ими по биологической очистке водоемов, представляется поистине грандиозной.
В пресных водах активными биофильтраторами являются перловицы (Unioninae), беззубки (Anodontinae), дрейссены (Dreissena), дафнии (Daphnia) и другие беспозвоночные. Значение их как своеобразной биологической «очистительной системы» водоемов настолько велико, что переоценить его практически невозможно.
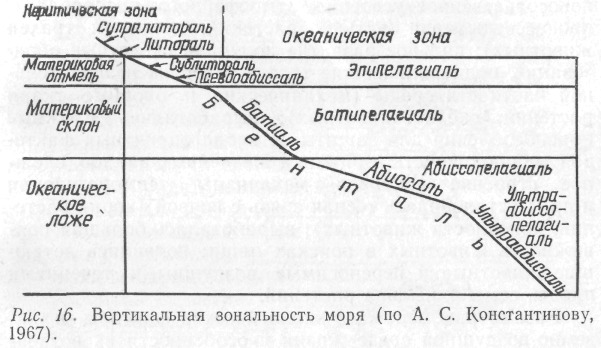
Зональность водной среды. Для водной среды жизни характерна четко выраженная горизонтальная и особенно вертикальная зональность. Все гидробионты строго приурочены к обитанию в определенных зонах, различающихся различными условиями обитания.
В Мировом океане толща воды носит название пелагиаль, а дно – бенталь. Соответственно различают и экологические группы организмов, обитающих в толще воды (пелагические) и на дне (бентосные).
Дно в зависимости от глубины его залегания от поверхности воды, делится на сублитораль (область плавного понижения до глубины 200 м), батиаль (крутой склон), абиссаль (океаническое ложе со средней глубиной 3–6 км), ультраабиссаль (дно океанических впадин, расположенное на глубине от 6 до 10 км). Выделяется еще литораль – кромка берега, периодически заливаемая во время приливов (рис. 16).
Открытые воды Мирового океана (пелагиаль) также делятся на зоны по вертикали соответственно зонам бентали: эпипелагиаль, батипелагиаль, абиссопелагиаль.
Наиболее богато растениями и животными заселены литоральная и сублиторальная зоны. Здесь много солнечного света, низкое давление, значительные колебания температуры. Обитатели абиссальных и ультраабиссальных глубин живут при постоянной температуре, в темноте, испытывают громадное давление, достигающее в океанических впадинах нескольких сотен атмосфер.
Подобная, но менее четко выраженная зональность характерна и для внутренних пресных водоемов.
Наземно-воздушная среда
Общая характеристика. Особенностью наземно-воздушной среды жизни является то, что организмы, обитающие здесь, окружены воздухом – газообразной средой, характеризующейся низкими влажностью, плотностью и давлением, а также высоким содержанием кислорода. Абсолютное большинство животных в этой среде передвигается по твердому субстрату – почве, а растения укореняются в ней.
Действующие в наземно-воздушной среде экологические факторы отличаются рядом специфических особенностей: свет здесь сравнительно с другими средами интенсивнее, температура претерпевает более сильные колебания, влажность значительно изменяется в зависимости от географического положения, сезона и времени суток. Воздействие почти всех этих факторов тесно связано с движением воздушных масс – ветра.
У обитателей наземно-воздушной среды в процессе эволюции выработались специфические анатомо-морфологические, физиологические, поведенческие и другие адаптации. У них появились органы, обеспечивающие непосредственное усвоение атмосферного кислорода в процессе дыхания (устьица растений, легкие и трахеи животных); сильное развитие получили скелетные образования, поддерживающие тело в условиях незначительной плотности среды (механические и опорные ткани растений, скелет животных); выработались сложные приспособления для защиты от неблагоприятных факторов (периодичность и ритмика жизненных циклов, сложное строение покровов, механизмы терморегуляции и др.); установилась тесная связь с почвой (корни растений, конечности животных); выработалась большая подвижность животных в поисках пищи; появились летающие животные и переносимые воздушными течениями плоды, семена, пыльца растений.
Рассмотрим основные экологические факторы в наземно-воздушной среде жизни и особенности их воздействия на наземные растения и животных.
Воздух. Воздух как экологический фактор характеризуется постоянством состава – кислорода в нем обычно около 21 %, углекислого газа 0,03 %. Без воздуха не могут существовать ни зеленые растения, ни аэробные микроорганизмы, ни животные. Кислород необходим для дыхания абсолютного большинства организмов, а углекислый газ используется при фотосинтезе.
Незначительная плотность воздушной среды не оказывает существенного сопротивления организмам при их передвижении по поверхности земли и в то же время затрудняет перемещение по вертикали. В процессе эволюции лишь немногие организмы поднялись в воздух и приспособились к полету (насекомые, птицы, из млекопитающих рукокрылые).
Воздух, как и другие факторы среды, оказывает на организмы прямое и косвенное действие. При прямом воздействии он имеет небольшое экологическое значение. Косвенное влияние воздуха осуществляется через ветры, которые, кроме того, что меняют характер таких важных факторов, как температура и влажность, оказывают механическое действие на организмы. Нередко сильные ветры, дующие в одном направлении, изгибают ветви и стволы деревьев в подветренную сторону, что служит причиной появления флагообразных форм кроны (рис. 17).

Определенную роль играет ветер в опылении растений-анемофилов (ветроопыляемые растения), которые выработали для этого ряд приспособлений: цветочные покровы у них обычно редуцированы и пыльники ничем не защищены от ветра.
Восходящие и особенно нисходящие токи нередко создают условия для застаивания и накопления у поверхности почвы холодного воздуха. Это вызывает задержку в развитии и растений и животных.
В районах, где постоянно дует сильный ветер, обычно беден видовой состав мелких летающих животных, поскольку они не могут противостоять ему при полете. У животных, обитающих в таких местах, развиваются плотные покровы, предохраняющие тело от охлаждения и потерь влаги. Птицы, живущие на открытых пространствах и в затишных местах, несколько различаются по строению. Это касается и насекомых. Так, на океанических островах с постоянными сильными ветрами многие птицы и особенно насекомые утрачивают крылья и способность к полету.
Воздушные потоки выполняют определенную роль и в расселении растений и животных. Плоды растений-анемохоров имеют множество приспособлений, увеличивающих их парусность, и разносятся ветром на громадные расстояния. Это различные хохолки, крылатки, парашюты, мешковидные полые вздутия. На большие расстояния переносятся ветром споры микроорганизмов, цисты простейших. Даже более крупные животные используют потоки воздуха для расселения. Пауки разносятся ветром на паутине, активно летающие насекомые – постоянными токами воздуха на больших высотах. Известны случаи заноса птиц сильными ветрами на значительное расстояние от их обычных мест обитания. Помимо случайного переноса животных ветром, многие виды активно используют воздушные течения для расселения. Ветры определяют направление миграций таких насекомых, как луговой мотылек (Loxostege stictialis), пустынная саранча (Schistocerca gregaria), малярийные комары (Anophe-· les).
Атмосферные осадки. Количество осадков, их распределение в течение года, форма, в которой они выпадают, в той или иной степени влияют на водный режим среды. Осадки в виде дождя, града или снега изменяют влажность воздуха и почвы, обеспечивают доступной влагой растения, дают питьевую воду животным. Сильные дожди могут временно затопить ту или иную территорию, вызвать паводки. Ливни и особенно град нередко приводят к механическому повреждению вегетативных органов растений.
Большое значение для водного режима имеют сроки выпадения дождей, их частота и продолжительность. Обилие дождей в период похолоданий практически не дает растениям необходимой влаги. Летом же при сравнительно высокой температуре даже незначительные осадки обеспечивают их нужным количеством воды.
Важен также характер дождей. При ливневых дождях почва не успевает впитывать воду. Эта вода сильными потоками быстро стекает и нередко сносит в реки и озера часть плодородного слоя почвы, а вместе с ней и слабо укоренившиеся растения, мелких животных. Моросящие дожди, наоборот, хорошо увлажняют почву и создают наиболее благоприятные условия для растений и животных. Но если они затягиваются, наступает переувлажнение. Это отрицательно сказывается на животных (на их размножении), приводит к сокращению их численности.
В поймах рек дожди могут вызвать паводки и оказать тем самым крайне неблагоприятное воздействие на обитающих здесь животных и на растения. В периодически затопляемых местах формируется своеобразная пойменная флора и фауна.
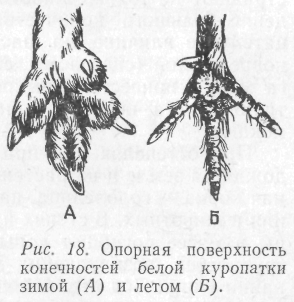
Значение снежного покрова было детально изучено А. Н. Формозовым (1946). Он показал, что снег выступает как хороший изолятор, защищающий почву и растительность от промерзания (слой снега в 20 см защищает растение при температуре воздуха – 25 °С), а для мелких животных служит укрытием, где они проводят значительную часть жизни, находя там благоприятные пищевые и температурные условия. При морозах свыше–14 °С под слоем снега в 20 см температура почвы не опускается ниже 0,2 °С, и в многоснежные зимы при наличии витаминизированного корма там могут размножаться грызуны. Под снегом при сильных морозах прячутся тетерева (Lyrurus tetrix), куропатки (Perdix perdix), рябчики (Tetrastes bonasia). Однако при многоснежных зимах наблюдается массовая гибель косуль (Capreolus capreolus), диких кабанов (Sus scrofa). Многим животным при мощном снежном покрове трудно передвигаться и добывать корм. И если лоси (Alces alces), например, свободно преодолевают слой снега глубиной до 50 см, то более мелким животным это недоступно.
Вместе с тем животные приспосабливаются к зимним условиям. К зиме у некоторых из них увеличивается опорная поверхность ног в результате обрастания их жесткими волосами, перьями, роговыми щитками (рис. 18); другие либо мигрируют, либо впадают в неактивное состояние (сон, спячка, диапауза); ряд животных переходит на питание определенными кормами. Однако, когда снега выпадает мало, гибнут кроты, мышевидные грызуны и другие мелкие животные.
В малоснежные зимы при сильных ветрах, когда снег сдувается с открытых пространств, от низких температур страдают не только животные, но и растения. Но выпадение большого количества снега оказывает и отрицательное влияние на растения. Кроме механических повреждений (снеголомы, снеговалы), мощный слой снега может привести к выпреванию растений. Во время таяния снега, особенно в затяжную весну, растения вымокают.
При оттепелях или при резком похолодании после дождя на земле и на растениях нередко образуется ледяная корка – гололедица, пагубно действующая на растения и животных. В степях и пустынях во время гололедицы в массе погибают копытные и многие птицы из-за трудностей, возникающих при передвижении и добывании пищи. Под ледяной коркой часто гибнут от удушения и вымерзания посевы культурных растений и дикорастущие травы.
При оттепелях на поверхности снега образуется наст – уплотняется снеговой покров. Это уменьшает его защитное свойство, затрудняет жизнь многих животных, передвигающихся под снегом и добывающих там корм. Ледяная корка на снегу может, как и гололедица, вызвать удушение растений, поскольку газообмен с наружным воздухом затрудняется, а расход кислорода растениями, микроорганизмами и животными, живущими под снегом, не прекращается.
В умеренных широтах, где зимой выпадают осадки в виде снега, животные и растения исторически приспособились к жизни в снегу или на его поверхности, выработав различные анатомо-морфологические, физиологические, поведенческие и другие особенности.
Различные осадки, помимо непосредственного воздействия на организмы, обусловливают ту или иную влажность воздуха, которая, как отмечалось, играет важную роль в жизни животных и растений, поскольку влияет на интенсивность их водного обмена. Чем меньше воздух насыщен парами воды, тем интенсивнее идут испарения с поверхности тела животных и транспирация у растений. Многие растения имеют специальные устьица, которые выделяют воду, когда воздух насыщен или близок к насыщению парами и обычная транспирация затруднена. Так животные и растения приспосабливаются к сезонному распределению осадков, к их количеству и характеру. Это определяет состав растительного и животного населения, сроки протекания тех или иных фаз в цикле их развития.
На влажность оказывает влияние и конденсация водяных паров, которая часто происходит в приземном слое воздуха при смене температуры. Проявляется это в выпадении росы при снижении температуры в вечерние часы. Роса нередко осаждается в таком количестве, что обильно смачивает растения, стекает в почву, увеличивает влажность воздуха и создает благоприятные условия для живых организмов, особенно когда других осадков выпадает мало. Осаждению росы способствуют растения: охлаждаясь ночью, они конденсируют на себе водяные пары. На режим влажности значительно влияют также туманы, густая облачность и другие природные явления.
Влажность почв. Одним из основных источников влаги для растений является почвенная вода. Однако не вся вода, содержащаяся в почве, может быть использована растениями. По физическому состоянию, подвижности, доступности и значению для растений почвенная вода подразделяется на свободную, капиллярную, химически и физически связанную.
Основной разновидностью свободной воды является гравитационная вода. Она заполняет широкие промежутки между частицами почвы и под действием силы тяжести постоянно уходит в более глубокие слои. Растения легко усваивают ее, пока она находится в зоне корневой системы. Запасы гравитационной воды пополняются осадками. Поэтому для растений весьма важен полив почвы, смачивание ее водой.
При сильных или продолжительных осадках образуется поверхностная свободная вода, поскольку ее приток значительно превышает скорость впитывания в почву. Такая вода стекает по уклону местности. Хотя свободная вода легко усваивается растениями, она вызывает временное или постоянное избыточное увлажнение, а это обусловливает заболачивание почвы и анаэробные процессы, снижающие продуктивность растений.
Капиллярная вода заполняет тончайшие промежутки между частицами почвы – капилляры. Она называется также подвешенной, поскольку не перемещается вниз под действием силы тяжести, а удерживается в капиллярах силой сцепления. Под влиянием испарения с поверхности почвы капиллярная вода образует восходящий ток в отличие от гравитационной, которой свойствен нисходящий ток. Вообще же капиллярная влага направляется в сторону меньшей влажности и поэтому может распространяться и горизонтально (подобно рассасыванию капли воды на фильтровальной бумаге). Капиллярная влага хорошо усваивается растениями.
И капиллярная и гравитационная воды почвы представляют собой так называемую доступную для растений воду.
Но в почве еще есть химически и физически связанная вода. Многие минералы почв (гипс, мирабилит, хлорид магния и др.) содержат до 50–60 весовых процентов воды. Вода входит в состав кристаллической решетки вторичных глинных минералов (кристаллизационная вода). Химически связанная и кристаллизационная вода неподвижна и совершенно недоступна растениям.
Физически связанная вода по степени подвижности образует две формы: пленочную, или рыхлосвязанную, и гигроскопическую, или прочносвязанную.
Пленочная вода представлена сотнями рядов диполей, последовательно облекающих друг друга. Сила, с которой она удерживается на поверхности почвенных частиц, не превышает 1 –10 атм. Пленочная вода способна передвигаться в почве в сторону меньшей влажности и ограниченно доступна растениям. Осмотическое давление клеточного сока корней позволяет им всасывать эту воду.
Гигроскопическая вода окутывает частицы почвы тонкой пленкой, и удерживается у почвенных частиц давлением 10 000–20 000 атм. Гигроскопическая вода высвобождается только при 105–110°С и физиологически совершенно недоступна растениям. Она образует так называемый мертвый запас воды в почве.
Количество недоступной растениям воды зависит от механического состава и физических свойств почвы, количества и качества органического вещества в ней, сосущей силы корней, относительной влажности воздуха. В песчаных почвах такая вода составляет 1–2 %, а в глинистых и торфяных достигает 50 % от общего количества воды. Когда в почве остается только недоступная вода, растение вянет и погибает.
Содержание влаги в почве, при котором растение не удовлетворяет свою потребность в воде, называется коэффициентом завядания. Для одного и того же вида растений на разных почвах коэффициент завядания неодинаков и составляет, например, для крупного песка 0,9 %, а для тяжелой глины– 16,3 %.
Таким образом, общее количество воды в почве не может характеризовать степень обеспеченности растений влагой. Чтобы определить ее, из общего количества воды необходимо вычесть коэффициент завядания.
Вместе с тем физически доступная вода почвы физиологически не всегда доступна растению. Низкая температура почвы, недостаток кислорода в почвенной воде и почвенном воздухе, кислотность почвы, высокая концентрация растворенных в почвенной воде минеральных солей – все это затрудняет усвоение почвенной влаги растением. Иногда летом после прохладной дождливой ночи многие растения проявляют явные признаки завядания, хотя почва сильно увлажнена и температура воздуха довольно высока. Причиной этого является низкая температура почвы, отрицательно действующая на всасывающую способность корней, и теплый воздух, обусловливающий довольно интенсивную транспирацию в надземных частях растения. В результате такого несоответствия между всасыванием воды корнями и отдачей ее листьями и происходит завядание растительности.
От количества физиологически доступной воды зависит развитие и надземных частей, и корневой системы растений. Как правило, у растений, произрастающих на сухих почвах, корневая система бывает более разветвленной, более мощной, чем на влажных.
Одним из источников почвенной влаги являются грунтовые воды. При низком их уровне капиллярная вода не достигает почвы и не влияет на ее водный режим. Увлажнение же почвы только атмосферными осадками вызывает сильные колебания ее влажности, что часто отрицательно влияет на растения. Вредно сказывается и слишком высокий уровень грунтовых вод, так как это приводит к переувлажнению почвы, к обеднению ее кислородом и обогащению минеральными солями. Оптимальный уровень грунтовых вод обеспечивает постоянное увлажнение почвы независимо от капризов погоды.
Итак, почвенная вода находится в постоянном движении: нисходящий ток гравитационной воды в сухие периоды сменяется восходящим током капиллярной воды. Эти движения воды, ее расход зависят от температуры, особенностей рельефа, свойств почвы, растительного покрова, перемещения воздуха и многих других факторов.
Экоклимат и микроклимат. Для того чтобы выяснить влияние климатических факторов на организм, часто метеорологических данных бывает недостаточно. Хорошо известно, что поверхности предметов, обращенные к солнцу, всегда теплее, чем воздух над ними; холодный воздух по ночам скапливается в понижениях местности. В связи с этим различные местообитания организмов отличаются температурным и световым режимом, режимом влажности. Иными словами, каждое местообитание характеризуется определенным экологическим климатом– экоклиматом, т. е. климатом приземного слоя воздуха.
Большое влияние на климатические факторы оказывает растительность. Под пологом леса, например, влажность воздуха всегда выше, а колебания температуры меньше, чем на полянах. Различен и световой режим этих мест. В разных растительных ассоциациях формируется свой режим влажности, температуры, света. Тогда говорят о фитоклимате.
Но не всегда данных экоклимата или фитоклимата достаточно для полной характеристики климатических условий того или иного местообитания. Условия жизни, окружающие личинок насекомых, живущих под корой дерева, иные, чем в лесу, где это дерево произрастает. При этом температура южной стороны ствола может быть на 10–15 °С выше температуры его северной стороны. Такие небольшие участки местообитания имеют свой микроклимат.
Четких различий между экоклиматом и микроклиматом не существует. Считается, что экоклимат – это климат относительно больших территорий, а микроклимат – климат отдельных небольших участков.
Каждая климатическая зона отличается огромным количеством разнообразных микроклиматов. Микроклиматы тесно связаны с солнечной радиацией, силой и направлением ветра, особенностями рельефа, характером растительности и другими климатическими показателями местности (рис. 19).
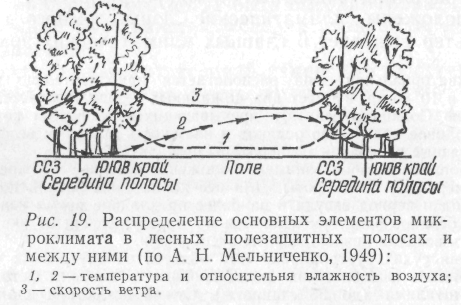
Особые микроклиматические условия создают не только растения, но и животные. Устойчивым микроклиматом обладают заселенные животными норы, дупла деревьев, пещеры. Наличие в одной местности многих микроклиматов обеспечивает сосуществование видов, обладающих неодинаковыми требованиями к внешней среде.
Географическая зональность и вертикальная поясность. Для наземно-воздушной среды, также как и для водной, характерна четко выраженная зональность. При этом любые сочетания растительного покрова и животного населения соответствуют морфологическим подразделениям географической оболочки Земли – климатическим зонам. Различают широтные и меридиональные, или долготные, природные зоны. Первые тянутся с запада на восток, вторые–с севера на юг. В долготном направлении широтные зоны подразделяются на подзоны, а в широтном – на провинции.
Животные и растения тесно связаны с условиями, характерными для каждой зоны, в которой они обитают, и адаптируются прежде всего к комплексу климатических факторов, которые в свою очередь зависят от почвенных условий, рельефа, от географических и ряда других особенностей той или иной местности. Вот почему в основу распределения биогеоценозов с их специфическим набором растений, животных и микроорганизмов могут быть положены климатические зоны земного шара. Г. Вальтер выделяет 6 главных климатических зон.
Экваториальная зона располагается приблизительно между 10°с. ш. и 10° ю. ш. и имеет два дождливых сезона (соответственно положению Солнца в зените), прерывающихся лишь на короткое время. Годовое количество осадков и влажность при этом весьма велики, месячные колебания температуры незначительны.
Тропическая зона занимает положение севернее и южнее первой, примерно до 30° широты. Для нее характерны летний дождливый период и период засухи в наиболее прохладное время года. Количество осадков здесь уменьшается по мере удаления от экватера; влажность воздуха низкая, морозы очень редки.
Зона сухих субтропиков – это области во внутренних частях субтропических океанических антициклонов со слабыми ветрами и частыми штилями (до 35° широты). Сумма осадков и влажность в этой зоне незначительны, годовые и суточные колебания температуры весьма существенны (бывают заморозки).
Переходная зона характеризуется сезонами зимних дождей. Морозы отмечаются чаще, чем в предыдущей зоне; лето жаркое. Зона охватывает Средиземноморье, Калифорнию, юг и юго-запад Австралии, юго-запад Южной Америки.
Умеренная зона отличается циклоническими дождями. Годовое количество осадков в ней уменьшается по мере удаления от океана; годовое колебание температуры становится более резким – лето жаркое, зима холодная. Зона подразделяется на следующие подзоны:
подзона теплого умеренного климата, где зимний период почти не выделяется и все времена года более или менее влажные (Южная Африка);
подзона типичного умеренного климата с холодной, но непродолжительной зимой, прохладным летом (Центральная Европа);
подзона аридного умеренного климата континентального типа с резкими температурными контрастами, небольшой суммой осадков и незначительной влажностью воздуха (Центральная Азия);
подзона бореального, или холодного, умеренного климата. Охватывает Северную Америку и северную часть Евразии. Лето здесь прохладное и влажное, зима длится более половины года.
6. Арктическая (антарктическая) зона. Здесь выпадает незначительное количество осадков преимущественно в виде снега. Лето (полярный день) короткое и прохладное. Зона переходит в полярную область, в которой произрастание растений невозможно.
Каждая климатическая зона характеризуется своеобразной растительностью и животным населением. Наиболее богаты жизнью и продуктивны тропические леса, поймы крупных рек, прерии и леса субтропиков и переходной зоны. Менее продуктивны пустыни, луга и степи.
Зональное распределение биогеоценозов обусловлено рядом факторов. Если, к примеру, то или иное растение или животное не встречается в какой-либо природной зоне, причиной этого могут быть высокие горные хребты или водные пространства, условия влажности, недостаток пищи и др.
Одним из важных условий изменчивости организмов и их зонального размещения по планете служит изменчивость химического состава среды. В этом отношении большое значение имеет учение А. П. Виноградова о биогеохимических провинциях, которые определяются зональностью химического состава почв, а также климатической, фитогеографической и геохимической зональностью биосферы. Биогеохимические провинции – это области на поверхности Земли, различающиеся по содержанию (в почвах, водах и т. д.) химических соединений, с которыми связаны определенные биологические реакции со стороны местной флоры и фауны.
Наряду с горизонтальной зональностью в наземной среде четко проявляется вертикальная поясность. В целом для планеты характерно изменение животного населения и растительности в направлении от основания горных областей к вершинам, подобно широтной зональности. При подъеме в горы повторяется та же смена зон, что и от экватора к полюсам. У подножия обычно располагаются пустыни, затем степи, широколиственные леса, хвойные леса, тундра и, наконец, льды. Однако полной аналогии все же нет. Горы обладают специфичной растительностью и животным населением.
Почва как среда жизни
Свойства почвы как экологического фактора (эдафические факторы). Почва как среда обитания обладает специфическими физическими свойствами. Для нее характерна более или менее рыхлая структура, определенная водопроницаемость и аэрируемость. Благодаря высокой дисперсности частиц почвы атмосферные осадки, как отмечалось, в виде дождя проникают в более глубокие слои ее и удерживаются там в капиллярных системах. А сами частицы несут на своей поверхности различные ионы, газы, пары воды.
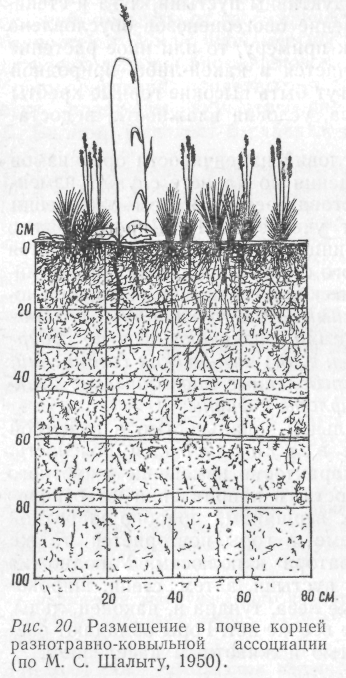
Почва обладает также своеобразными биологическими особенностями, поскольку тесно связана с жизнедеятельностью организмов. Верхние слои ее содержат массу корней растений. В процессе роста, отмирания и разложения они разрыхляют почву и создают определенную структуру ее, а вместе с тем и условия для жизни других организмов (рис. 20). Роющие животные перемешивают почвенную массу, а после смерти становятся источником органического вещества для микроорганизмов.
Все свойства почвы во многом зависят не только от климатических факторов, но и от жизнедеятельности почвенных организмов, которые механически перемешивают ее и перерабатывают химически, создавая в конечном итоге необходимые для себя условия. При участии организмов в почве происходит постоянный круговорот веществ и миграция энергии. Климатические факторы, а также жизнедеятельность растений, животных и микроорганизмов обусловливают ежегодные циклические изменения почвы, специфичные для разных широт.
Большую роль в формировании почвы играет рельеф. На одинаковых и одновозрастных формах рельефа образуются близкие или даже однотипные почвы. На местности с расчлененным рельефом, неодинаковым уровнем грунтовых вод наблюдаются различия в климате, режиме тепла и скорости испарения поверхностной влаги, а также в распределении атмосферных осадков. Все это существенно влияет на химические и физические свойства почв и вместе с тем на характер растительного покрова и животного населения.
Свойства почвы в своей совокупности создают определенный экологический режим ее, основными показателями которого служат гидротермические факторы и аэрация.
Хорошо увлажненная почва легко прогревается и медленно остывает. На поверхности ее происходят более резкие колебания температуры, чем в глубине. При этом суточные колебания ее затрагивают слои до глубины в 1 м. И если учесть, что зимой температура почвы с глубиной повышается, а летом, наоборот, падает, то легко можно представить сезонные вертикальные миграции почвенных обитателей, вызванные изменением условий среды. Естественно, зимой почвенные животные находятся глубже, чем летом.
Аэрацию почвы обусловливает ее пористость, которая обеспечивает циркуляцию не только воды, но и воздуха. На аэрацию почвы влияют различные факторы. Так, повышение влажности препятствует проникновению в почву кислорода. С глубиной количество углекислого газа в почве увеличивается, а кислорода уменьшается. При повышении температуры воздух расширяется и выходит из почвы, а при понижении он более интенсивно проникает в промежутки между почвенными частицами. Следовательно, соотношения воды и воздуха в почве весьма изменчивы. Это также является причиной вертикальных миграций почвенных организмов. При избытке влаги и ухудшении аэрации они поднимаются к поверхности, при недостатке влаги и улучшении аэрации – опускаются в более глубинные слои.
Гидротермический режим и аэрация почв зависят и от их характера. Например, поскольку скважность глинистых почв выражена слабее, чем песчаных, первые удерживают влагу сильнее и влажность их бывает выше. В то же время воздушный режим глинистых почв менее благоприятен для многих растений и животных. В результате глинистые почвы, как более влажные и менее воздухопроницаемые, прогреваются хуже, чем песчаные. Вот почему во влажном и прохладном климате песчаные почвы теплее глинистых. В сухом же и жарком поясе песчаные почвы более влажные. Поэтому в песчаных пустынях жизнь богаче, чем в глинистых,– растения достигают здесь более крупных размеров, а почвенные животные отличаются видовым разнообразием и многочисленностью.
Итак, влажность, температура и аэрация почвы - факторы тесно взаимосвязанные и взаимозависимые·. Сложный комплекс их обусловливает гидротермический режим почвы и в результате оказывает решающее влияние на существование почвенных обитателей.
Роль почвы в жизнедеятельности живых организмов. Благодаря специфическим свойствам почва выполняет одну из важных функций в жизни различных почвенных организмов и прежде всего растений – обеспечивает им водоснабжение и минеральное питание.
Оптимальные запасы доступной для растения почвенной воды – чрезвычайно существенный фактор. В почве различают биологически полезную и биологически бесполезную воду. Биологически полезной является вода, свободно передвигающаяся по капиллярам почвы и бесперебойно снабжающая растения влагой. Значение почвы в водоснабжении растений тем выше, чем она легче отдает им воду. Это зависит от структуры почвы и степени набухаемости ее частиц.
Различают физическую и физиологическую сухость почвы.
При физической сухости почва испытывает недостаток влаги. Происходит это при атмосферной засухе, когда поступление воды резко сокращается, что обычно наблюдается в сухом климате и в местах, где почва увлажняется только за счет атмосферных осадков.
Физиологическая сухость почвы - явление более сложное. Оно возникает в результате физиологической недоступности физически доступной воды. Растения при физиологической сухости страдают даже на влажных почвах, когда низкая температура почвенного покрова или другие неблагоприятные условия препятствуют нормальному функционированию корневой системы. Так, на сфагновых болотах, несмотря на большое количество влаги, вода оказывается недоступной для многих растений из-за высокой кислотности почвы, плохой аэрации ее и наличия токсических веществ, нарушающих нормальную физиологическую функцию корневой системы. В тундре из-за того что интенсивность поступления воды из глубоких и очень охлажденных слоев не соответствует интенсивности испарения, также отмечается физиологическая сухость почвы. Физиологически сухими являются и сильно засоленные почвы. Из-за высокого осмотического давления почвенного раствора вода засоленных почв для многих растений оказывается недоступной.
Почва играет важную роль ив минеральном питании растений. Вместе с водой в растения через корневую систему поступает ряд минеральных веществ, находящихся в почве в растворенном состоянии. Однако корневое питание растений – не простое всасывание веществ, а сложный биохимический процесс, в котором особую роль играют почвенные микроорганизмы. Известно, что в прикорневой зоне, в ризосфере, количество микроорганизмов всегда больше, чем на некотором расстоянии от нее. Это связано с тем, что корневые выделения растений оказывают положительное влияние на жизнедеятельность микроорганизмов, а выделения последних усваиваются корневой системой. Вот почему большинство высших растений имеют микоризу, значительно увеличивающую активную поверхность корней.
Органическое вещество почвы играет важную роль в росте и развитии растений. Оно состоит из продуктов гумификации (аэробное разложение) и неполного разложения растительных остатков и трупов почвенных животных. Перегной, или гумус, для всех почвенных обитателей является основным источником необходимых для жизни минеральных соединений и энергии. Он обусловливает плодородие почв и их структуру. Процессы минерализации свежих органических веществ и перегноя обеспечивают постоянное поступление в почвенный раствор таких важнейших элементов питания растений, как азот, фосфор, сера, кальций, калий, микроэлементы. Жизнедеятельность растений, образование гумуса и его минерализация превращают элементы минерального питания из почвообразующих пород в доступные для растений соединения и концентрируют их в верхних горизонтах почвы. Гумус служит источником физиологически активных соединений (витамины, органические кислоты, полифенолы и др.), которые стимулируют рост растений. Перегнойные вещества обеспечивают также водоустойчивую структуру почв, что создает благоприятный для растений водно-воздушный режим.
Главным энергетическим материалом в почве является органическое вещество корней. От его количества зависит численность и видовое разнообразие почвенных обитателей. Высокое содержание растительных остатков в почве способствует значительному развитию сапрофагов. Это и животные, и микроорганизмы, и бесхлорофилльные растения.
Животные-сапрофаги выполняют существенную роль в круговороте веществ в почве. Питаясь мертвыми органическими соединениями и обеспечивая пищей хищных почвенных животных, они служат важным звеном процесса превращения органического вещества в минеральное. Завершает процесс разложения растительного и животного органического вещества вплоть до полной минерализации микробное население почвы.
Распределение органического вещества в почве и плодородие последней находится в тесной зависимости от климатических условий, гидротермического режима почвы, ее аэрации и других факторов. При наиболее благоприятном сочетании этих факторов происходит гумификация органических остатков и тем самым создаются высокоплодородные почвы – черноземы, черные почвы тропиков, бурые лесные. В таких почвах развиваются богатые фауна и флора и интенсивно накапливается и разлагается органическое вещество.
Роль микроорганизмов, высших растений и животных в почвообразовательных процессах. Микроорганизмы, растения и животные, обитающие в почве, находятся в постоянном взаимодействии друг с другом, а также и со средой обитания. Эти взаимоотношения очень сложны и многообразны. Животные и бактерии потребляют растительные углеводы, жиры и белки. Грибы разрушают целлюлозу, в частности древесину. Хищники питаются живыми тканями своих жертв. Благодаря этим взаимоотношениям и в результате коренных изменений физических, химических и биохимических свойств горной породы в природе постоянно происходят почвообразовательные процессы.
Наиболее древними почвообразователями являются микроорганизмы. Они функционировали задолго до появления на Земле высших растений и животных. Микроорганизмы осуществляют в почвах и в целом в биосфере процессы, играющие важную роль в биологическом круговороте веществ и потоке энергии. Эти процессы недоступны высшим организмам. Синтез физиологически активных соединений, гумусообразование и полная минерализация органических остатков – вот главная функция микроорганизмов в почвообразовательных процессах.
В числе почвенных микроорганизмов имеются представители и растительного и животного мира. Прежде всего это одноклеточные и многоклеточные водоросли. Многие из них обладают пигментами типа хлорофилла и отличаются от других микроорганизмов тем, что способствуют обогащению почвы органическим веществом и кислородом. Однако наиболее многочисленны в почвенной микрофлоре грибы, актиномицеты и бактерии. В почвенной микрофауне особое значение имеют амебы, жгутиковые и ресничные, а также специфичные микронематоды. Есть сведения о присутствии в почве также неклеточных форм микроорганизмов – бактериофагов и вирусов. Они, являясь паразитами и хищниками бактерий, влияют на жизнедеятельность последних.
Основной функцией высших растений в почвообразовательном процессе служат синтез органического вещества и аккумуляция в нем солнечной энергии. Органическое вещество в процессе фотосинтеза накапливается в надземных и подземных частях растений, а после их отмирания переходит в почву и подвергается минерализации. Скорость процессов минерализации органического вещества и состав образующихся при этом соединений во· многом зависят от типа растительности. Продукты разложения хвои, листьев, древесины, травянистого покрова различны как по химизму, так и по влиянию на процесс почвообразования. В сочетании с другими факторами это приводит к формированию различных типов почв.
Главная функция животных в почвообразовательном процессе – это потребление и разрушение органического вещества, а также перераспределение запасов энергии.
В целом процесс гумусообразования начинается разрушением и измельчением растительной массы и мертвого животного вещества. Этот процесс осуществляется позвоночными животными при обязательном участии грибов и бактерий (рис. 21). К таким животным относятся фитофаги, питающиеся тканями живых растений; сапрофаги, потребляющие мертвое вещество растений; некрофаги, питающиеся трупами животных; хищники, поедающие живых животных; копрофаги, уничтожающие экскременты животных.
Большую роль в процессах почвообразования играют подвижные почвенные животные. Они разрыхляют почву, создают условия для ее аэрации, механически перемещают в почве органические и минеральные вещества. Например, дождевые черви на луговых почвах за год выбрасывают на поверхность до 80–90 т/га копрогенного материала, а степные грызуны перемещают вверх и вниз сотни кубических метров грунта и органического вещества.
Итак, круговорот веществ в почве можно представить следующим образом. Растения синтезируют органическое вещество. Животные производят механическое и биохимическое разрушение его и как бы подготавливают для гумусообразования. Микроорганизмы синтезируют почвенный гумус и затем разлагают его.
Экологические группы почвенных организмов. Количество организмов в почве огромно. Однако, как указывает М. С. Гиляров, в связи со сглаженностью экологических условий все они отличаются «выравненностью группового состава». Кроме того, для них характерна повторяемость в различных климатических зонах.
Почвенные животные могут быть сгруппированы по степени связи со средой обитания в три основные экологические группы.
Геобионты – постоянные обитатели почвы. Весь цикл их развития протекает в почвенной среде. Типичными представителями являются дождевые черви (Lymbricidae), многие первичнобескрылые насекомые (Apterygota).
Геофилы – животные, часть цикла развития которых, чаще одна из фаз, обязательно проходит в почве. К ним принадлежит большинство насекомых: саранчовые (Acridoidea), ряд жуков (Staphylinidae, Carabidae, Elateridae), комары-долгоножки (Tipulidae). Личинки их развиваются в почве, а во взрослом состоянии это типичные наземные обитатели. К геофилам принадлежат и такие насекомые, которые в почве находятся в фазе куколки.
Геоксены – животные, иногда посещающие почву для временного укрытия или убежища. Из насекомых к геоксенам относятся таракановые (Blattodea), многие полужесткокрылые (Hemiptera), некоторые развивающиеся вне почвы жуки, а также грызуны и другие млекопитающие, живущие в норах.
Однако эта классификация не отражает роли животных в почвообразовательных процессах, поскольку в каждой группе есть организмы как активно передвигающиеся и питающиеся в почве, так и пассивные, которые пребывают в почве в период отдельных фаз развития (личинки, куколки или яйца насекомых). Поэтому почвенных обитателей можно разделить на экологические группы исходя из их размеров и степени подвижности. Выделяют три такие группы.
Микробиотип, микробиота, – почвенные микроорганизмы, составляющие основное звено детритной пищевой цепи. Они представляют собой как бы промежуточное звено между растительными остатками и почвенными животными. Это прежде всего зеленые (Chlorophyta) и сине-зеленые (Cyanophyta) водоросли, бактерии (Bacteria), грибы (Fungi) и простейшие (Protozoa).
Мезобиотип, мезобиота,– совокупность сравнительно мелких легко извлекающихся из почвы подвижных животных. Сюда относятся почвенные нематоды (Nematoda), мелкие личинки насекомых, клещи (Oribatei), ногохвостки (Collembola) и др. Они весьма многочисленны – от десятков и сотен тысяч до миллионов особей на 1 кв. м почвы. Питаются в основном детритом и бактериями. Некоторые клещи и насекомые являются хищниками, а отдельные виды нематод паразитируют в корнях растений, иногда сильно повреждая их.
Макробиотип, макробиота,– крупные насекомые, дождевые черви и другие животные вплоть до роющих позвоночных. В этот тип включаются и корни растений. Наибольшее значение в данной группе имеют дождевые черви. Численность их может достигать до 300 на 1 кв. м. Вместе с другими животными они играют большую роль в перемешивании почвы. Сюда относятся и подвижные членистоногие, обитающие между почвой и подстилкой. Механически разрушая последнюю, они переводят ее в удобную для разложения микроорганизмами форму.
Особую группу представляют организмы, заселяющие сыпучие подвижные пески. Растения, адаптированные к таким местам обитания, называются псаммофитами, а животные – псаммофилами. Подвижные сыпучие пески широко представлены в песчаных пустынях. Мелкими массивами они встречаются в степях и в лесной зоне.
Растения сыпучих песков во всех климатических зонах имеют общие особенности морфологии и биологии. Это объясняется аналогичностью экологических условий. Образующиеся в песках барханы из-за постоянно дующих ветров беспрерывно перемещаются. В результате растения либо засыпаются толстым слоем песка, либо, наоборот, обнажаются вплоть до корневой системы. Семенам растений на подвижном песке трудно прорастать и укореняться. Водный режим в этих местах также в основном одинаков, поскольку для песков характерна большая проницаемость и ничтожная водоудерживающая сила, малая капиллярность и глубокое залегание грунтовых вод.
У растений, произрастающих на подвижных песках, исторически выработались своеобразные приспособления. Древесные и кустарниковые псаммофиты, к примеру, при засыпании их песком образуют придаточные корни. Придаточные почки и побеги развиваются на корнях, если растения обнажаются при выдувании песка. Таковы белый саксаул (Arthrophytum haloxylon), кандымы (Calligonum), песчаная акация (Ammodendron conollyi) и другие типичные пустынные растения. Некоторые псаммофиты спасаются от заноса песком быстрым ростом побегов, редукцией листьев. Это приспособление к тому, чтобы песок легко продувался и не задерживался. У псаммофитов развились специальные приспособления, увеличивающие летучесть и пружинистость плодов. Их плоды передвигаются вместе с движущимся песком и не засыпаются им. Псаммофиты легко переносят засуху благодаря различным приспособлениям. Это и песчаные чехлы на корнях, и опробковение корней, и сильное развитие боковых корней. Разреженная растительность пустыни в значительной степени обусловлена тем, что подземная часть их (корневая система) довольно густая и обычно плотно сомкнутая. Большинство псаммофитов безлистые или имеют четко выраженную ксероморфную листву (листья жесткие с сизым восковым налетом, узкие и свернутые или блестящие и кожистые). Это значительно сокращает транспирирующую поверхность.
Сыпучие пески встречаются и во влажном климате. Примером могут служить песчаные дюны по берегам северных морей и в Прибалтике, пески обсыхающего речного ложа по берегам крупных рек. На этих песках растут такие типичные псаммофиты, как ива-шелюга (Salix acutifolia), волоснец песчаный (Elymus arenarius), овсяница песчаная (Festuca arenaria). Большинство из них характеризуется ксероморфностью.
К специфичным условиям жизни в сыпучих подвижных песках приспособлены и многие животные. У позвоночных псаммофилов конечности часто устроены в форме своеобразных «песчаных лыж», облегчающих передвижение по рыхлому грунту. У ящериц, живущих на песке (Psammodromus, Phrynocephalus), на пальцах развиваются роговые гребни. У тонкопалого суслика (Spermophilopsis leptodactylus) и гребнепалого тушканчика (Paradipus ctenodactylus) пальцы покрыты длинными волосами и роговыми выростами. Птицы и млекопитающие песчаных пустынь способны преодолевать большие расстояния в поисках воды или длительное время обходиться без нее. Это, например, бегунки (Cursorius cursor), рябки (Pterocletes). Верблюды благополучно переносят отсутствие воды до 8–16 суток, обеспечивая организм необходимой влагой за счет окисления жиров. Другие животные получают воду вместе с пищей или запасают ее в период дождей, накапливая в мочевом пузыре, в подкожных тканях, в брюшной полости. Некоторые животные во время засухи прячутся в норы, зарываются в песок или впадают в летнюю спячку.
В подвижных песках живут многие членистоногие. Типичными псаммофилами являются мраморные хрущи из рода Polyphylla, личинки муравьиных львов (Myrmeleonidae) и скакунов (Cicindelinae), многие перепончатокрылые (Hymenoptera). Почвенные животные, обитающие в подвижных песках, обладают специфическими приспособлениями, обеспечивающими им передвижение в рыхлом грунте. Обычно это «минирующие» животные, которые раздвигают частицы песка. Сыпучие пески заселяются только типичными псаммофилами.
В тесной связи с изучением особенностей обитателей подвижных песков находится проблема закрепления последних. При перевыпасе на сухих песчаных почвах часто нарушается задерненность, что облегчает выдувание, перемещение песка при ветре. Подвижные пески надвигаются на соседнюю растительность, на посевы. Это явление наблюдается и при ошибках, допущенных во время обработки почвы, при проведении мелиоративных работ. Для закрепления подвижных песков используются растения-псаммофиты: ива-шелюга, кандымы и другие кустарники. После закрепления песка псаммофитами создаются благоприятные условия для произрастания других растений. Так, на закрепленных дюнах разрастаются сосновые леса, на степных и пустынных песках успешно возделываются бахчевые, зерновые, кормовые и плодовые растения. При этом сами псаммофиты по мере уплотнения песка находят для себя все меньше благоприятных условий и в конце концов исчезают, что приводит к полной смене растительных ассоциаций, а вместе с тем и животного населения.
Засоленные почвы. В зонах недостаточного атмосферного увлажнения, в основном в степях и пустынях, широко распространены засоленные почвы. Это связано с тем, что в засушливом и жарком климате наблюдается неполное промывание почв осадками. В таких почвах преобладает восходящий ток воды, который приносит в верхние горизонты большое количество легкорастворимых солей, вредных для большинства растений и животных. Засоление почв может происходить и на низких побережьях морей и океанов, в местах выхода соленых источников и ключей.
Среди засоленных почв различают солончаки – почвы, испытывающие постоянное и сильное увлажнение солеными водами. При высыхании на их поверхности образуется корочка из кристаллических солей. В нашей стране солончаки отмечаются в Казахстане и Средней Азии, по берегам соленых озер.
Так называемые солонцы содержат вредные соли только в глубоких слоях, а на поверхности они сильно выщелочены.
Промежуточное положение между солончаками и незасоленными почвами занимают солончаковатые почвы. Они увлажняются сравнительно слабозасоленными водами.
Выделяются еще солончаковые солонцы и солонцеватые почвы. У первых поверхностные горизонты еще не окончательно выщелочены. Эти почвы занимают промежуточное положение между солончаками и солонцами. Солонцеватые почвы слабо засолены в нижних горизонтах и весной при определенном количестве осадков могут опресняться.
Растения, обитающие на засоленных почвах, называются галофитами. Все они имеют очень высокое осмотическое давление, позволяющее им использовать почвенные растворы, поскольку сосущая сила корней превосходит сосущую силу почвенного раствора. Некоторые галофиты выделяют излишки солей через листья или накапливают их в своем организме. Вот почему их иногда используют для получения соды и поташа. Типичными галофитами являются солерос (Salicornia herbacea), cap-сазан (Halocnemum strobilaceum), содовая (Salsola soda) и калийная (S. kali) солянки, поташник (Kalidium). Многие галофиты охотно поедаются животными и в некоторых местах служат для них основным кормом в течение круглого года.
Имеются и животные, приспособившиеся к жизни на засоленных почвах. Это так называемые галофилы.
В целом почвенные животные относительно солевыносливы, поскольку концентрация солей в почвах может существенно колебаться в связи с периодической сменой засух дождями. Однако в засоленных почвах фауна и в количественном и в качественном отношении сильно обедняется – исчезают, к примеру, личинки щелкунов (Elateridae), хрущей (Melolonthinae), но зато появляются специфичные галофилы, не встречающиеся в почвах обычной засоленности. К ним можно отнести личинок некоторых пустынных жуков-чернотелок (Tenebrionidae).
Весьма своеобразные экологические условия складываются на меловых обнажениях: резко выраженный специфичный химизм, каменистость, особый световой режим в связи с сильным отражением света (альбедо).
На южных склонах меловых обнажений встречаются южные ксерофиты, на северных – арктические и альпийские растения. Подземные органы у меловых растений часто деревянистые или, наоборот, мясистые и сочные. Листья опушенные или свернутые, обычно небольшие. У многих хлоропласта расположены в эпидермисе. Для некоторых характерно сравнительно высокое осмотическое давление (до 25–29 атм). Заселяют такие растения площадки мела, свободные от других растений, что свидетельствует о их высокой выносливости в крайне неблагоприятных условиях.
О животных, приспособленных к жизни на меловых обнажениях, имеется очень мало сведений, а экология типичных меловых обитателей практически не изучена.
Значение эдафических факторов в распределении растений и животных. Как отмечалось, в связи с разнообразием условий мест обитания, в том числе и почвенных, а также в связи с избирательностью по отношению к ним растений в определенной ландшафтно-географической зоне формируются специфические растительные ассоциации. При этом даже в одной зоне в зависимости от ее рельефа, уровня грунтовых вод, экспозиции склона и ряда других факторов создаются неодинаковые почвенные условия, что отражается на типе растительности. Например, в ковыльно-типчаковой степи всегда можно обнаружить участки, где доминирует типчак или, наоборот, ковыль. Таким образом, типы почв являются мощным фактором распределения растений.
В меньшей степени эдафические факторы непосредственно влияют па наземных животных. Однако последние тесно связаны с растительностью и она играет решающую роль в их распределении. Тем не менее и среди крупных позвоночных легко обнаружить формы, приспособленные к конкретным почвам. Это особенно характерно для фауны сыпучих песков, глинистых почв с твердой поверхностью. Специфична фауна торфяников и заболоченных земель. В еще более тесной связи с почвенными условиями находятся роющие формы. Одни из них могут разрывать только легкие песчаные почвы, другие приспособлены к более плотным почвам.
Типичные почвенные животные также адаптированы к различным видам почв. К солонцам и солончакам, например, строго приурочены группы беспозвоночных-гало-Филов. (В Средней Европе известно до 20 родов жуков, распространенных только на солончаковых или солонцовых почвах.) Однако многие почвенные животные имеют очень широкие ареалы и встречаются в разных почвах. Так, дождевой червь (Eisenia nordenskioldi) достигает высокой численности в таежных и тундровых почвах, в почвах лугов и смешанных лесов, встречается даже в горах. Это объясняется тем, что в распространении почвенных обитателей большое значение, кроме свойств самой почвы, имеет их эволюционный уровень, а также размеры тела. Тенденция к космополитизму особенно выражена у мелких форм. Таковы бактерии, грибы, простейшие, микроартроподы (клещи, коллемболы), почвенные нематоды.
Итак, почва как среда обитания сформировалась в результате жизнедеятельности живых организмов. Возникнув, она заняла как бы промежуточное положение между водной средой и наземной и стала играть важную роль в дальнейшей эволюции животного мира. К примеру, многие группы членистоногих в процессе исторического развития прошли сложный путь от типично водных организмов через почвенных обитателей до типично наземных форм.
Живые организмы как среда жизни
Топические связи. Роль организмов в создании среды друг для друга. Понятие о топических связях ввел В. К. Беклемишев, подразумевая под ними воздействие одних организмов на другие через изменение различных абиотических факторов (движение воды и воздуха, освещение, температура, влажность). Результатом топических связей, по выражению Беклемишева, является «кондиционирование» среды, т. е. создание одним организмом определенных физических или химических условий для другого.
Топические связи играют важную роль в формировании биоценозов и могут быть благоприятными или неблагоприятными. Они оказывают существенное влияние на размещение организмов в сообществе. Под пологом леса, например, многие животные укрываются от непогоды, врагов, находят там определенные условия микроклимата, способствующие размножению и выкармливанию потомства. В лесных массивах благодаря растительности, как правило, повышается влажность воздуха, уменьшается амплитуда температурных колебаний в течение суток, гасятся движения воздуха, создается своеобразный химизм и температурный режим почвы, т. е. формируются условия абиотической среды, благоприятные для жизнедеятельности различных организмов. Лесной климат отличается большой устойчивостью метеорологических факторов, несмотря на то что средняя годовая температура и количество осадков в лесу могут мало отличаться от таковых на безлесных территориях.
На растениях находят убежища самые различные животные. Они поселяются в расщелинах коры, в листве, строят гнезда на ветвях. Своеобразную среду обитания предоставляют древесные растения вьющимся, лазающим растениям и эпифитам. С другой стороны, сами эпифиты приятные для жизнедеятельности различных организмов. В лесах Малакки, например, произрастает папоротник-эпифит олений рог (Platycerium grande). В розетках листьев его накапливаются опавшие листья и влага и со временем образуется гумус, который заселяется специфической фауной. Там можно обнаружить даже дождевых червей. У бромелиевых (Bromeliaceae) основания листьев расширены в виде своеобразных чаш. Здесь также собирается дождевая вода. Эти искусственные бассейны заселяются рядом организмов. В тропических лесах Южной Америки встречаются водные насекомые, обитающие только в таких «чашах». Здесь же в массе размножаются и малярийные комары.
Таким образом, в биоценозах организмы объединяются благодаря не только трофическим, но и топическим связям, которые часто тесно переплетаются, создавая в сообществе сложные межвидовые взаимоотношения.
Живые организмы как среда обитания. Для животных и растений, ведущих паразитический образ жизни, организм, на котором или в котором они поселяются (хозяин), является специфической средой обитания. Наиболее детально и всесторонне это явление описали В. А. Догель, Ε. Η. Павловский и ряд других исследователей. Они показали, что организм хозяина для паразита – это специфическая среда жизни.
Многие паразиты почти полностью утратили связь с внешним миром – все стадии их развития проходят в организме хозяев (малярийный плазмодий, трихина спиральная и др.). Между паразитами и хозяевами в процессе эволюции возникли сложные взаимоотношения. Поскольку любой организм зависит от условий среды и сам на нее воздействует, паразит не только зависит от хозяина, но и влияет на него. У хозяина в результате вырабатываются самые различные защитные реакции. Паразиты же приспосабливаются к этим реакциям, и, таким образом, процесс коадаптации происходит постоянно.
Большое разнообразие подобных взаимоотношений, различные их оттенки отражают пути возникновения паразитизма.
Первый путь – это простое «квартирантство», когда более мелкий организм поселяется в жилище более крупного или вблизи него и со временем переходит на тело хозяина, а затем и внутрь, переключаясь на питание за счет его пищи или соков и тем самым причиняя ему вред. Так квартирант может превратиться в паразита, и тело хозяина становится для него средой обитания.
Второй путь перехода к паразитизму – через хищничество. Если хищник нападает на крупную добычу, которую не может уничтожить и съесть сразу, он прикрепляется к ней и постоянно питается тканями или соками ее тела. При определенных условиях такой хищник проникает внутрь тела хозяина и, найдя там благоприятную среду – обилие пищи, превращается в паразита. И в этом случае организм хозяина становится средой обитания.
Третий путь – случайное проникновение будущего паразита в организм хозяина. Крупные животные могут заглатывать с пищей мелкие формы, некоторые из них не погибают, а, приспосабливаясь к новым условиям, превращаются в паразитов.
В природе сохранилось немало примеров этих трех путей перехода от квартирантства, хищничества и случайного паразитирования к подлинному паразитизму. Однако трудно сказать, когда на Земле появился первый паразит и от какой группы животных или растений он произошел.
Все паразиты делятся на две группы: эктопаразиты – наружные паразиты, обитающие на поверхности тела хозяина (клещи, пиявки, блохи), и эндопаразиты – внутренние паразиты, живущие внутри тела хозяина (большинство гельминтов, бактерий, вирусов, паразитические простейшие).
Кроме того, отличаются стационарный паразитизм – паразит на длительное время, часто на всю жизнь, связывает себя с хозяином. Стационарные паразиты могут быть постоянными (приурочены к одному хозяину – вши, пухоеды, чесоточные зудни и периодическими (развитие их протекает со сменой хозяев – многие ленточные черви, сосальщики). В простейшем случае они развиваются в окончательном и промежуточном хозяевах. Малярийный плазмодий, к примеру, определенную часть жизни проводит в малярийном комаре – окончательный хозяин. Промежуточным хозяином его является человек. Окончательным хозяином служит тот организм, в котором обитает половозрелая форма паразита, а промежуточным,– в котором паразит проходит личиночную, неполовозрелую стадию.
Существуют и временные паразиты. Они не всю жизнь связывают с хозяином, а часть ее проводят свободно (кровососущие двукрылые, многие клопы).
Паразиты бывают и факультативными. Для них паразитизм не обязателен. Так, миноги могут паразитировать на крупных рыбах либо вести хищнический образ жизни, поедая мелких животных. Облигатные же, обязательные, паразиты без хозяина существовать не могут.
И у животных, и у растений, ведущих паразитический образ жизни, выработались многочисленные приспособления к паразитированию в виде анатомо-морфологических и физиологических особенностей. Любой организм в естественных условиях заражен теми или иными паразитами – вирусами, бактериями, самыми разнообразными растениями и животными, приспособившимися в процессе эволюции к паразитированию. Нередко в одном хозяине поселяются несколько видов паразитов, локализуясь в различных органах и тканях.
Одна из форм приспособления к паразитизму – упрощение организации паразита по сравнению со свободно живущими предками. Примеры упрощения, доведенного до крайности, можно встретить и в растительном и в животном мире. Так, у ряда паразитических червей из класса ленточных (Cestoidea) редуцируются органы пищеварения и чувств в связи с тем, что они буквально «купаются» в пищевом бульоне и всасывают пищу всей поверхностью тела. Многие паразиты из насекомых утрачивают крылья (вши, блохи). Среди животных известен паразит из ракообразных – саккулина (Sacculina carcini). Тело ее состоит из мантии, половых желез и упрощенной нервной системы. Саккулина паразитирует на крабах и имеет вид небольшого мешочка, располагающегося на нижней стороне брюшка хозяина. От этого мешочка внутрь тела краба, наподобие корней, идут тонкие выросты, пронизывающие органы и ткани хозяина. У некоторых растений-паразитов сокращается количество зеленых частей тела, а у отдельных хлорофиллоносные органы исчезают полностью. В мире цветковых растений крайняя степень упрощения наблюдается у раффлезии (Rafflesia arnoldii) –паразита лианы циссус (Cissus). У нее сохранился лишь цветок, а все остальные органы превратились в нити. Последние внедряются в промежутки между клетками хозяина и напоминают мицелий гриба. Раффлезия и саккулина – примеры поразительного сходства между растением-паразитом и животным-паразитом.

Наряду с упрощением организации почти у всех паразитов, появляются специфичные органы фиксации (крючки, зацепки, присоски и др.). Они позволяют им прикрепляться и удерживаться на теле или в органах и полостях хозяина. В связи с необходимостью обязательного контакта с хозяином у паразитов выработались приспособления, обеспечивающие выживание вида даже в условиях, если такая встреча затруднена. Так, колдунья-трава (Striga), распространенная в Африке, Южной Азии и Австралии и поражающая корневую систему многих злаков, продуцирует до 500 000 семян, которые способны пролежать в земле, не прорастая, до 20 лет. Но как только рядом оказывается растение-хозяин, семя стриги трогается в рост и проникает в его корень. Семена заразихи также прорастают только под влиянием корневых выделений растения-хозяина.
Чрезвычайно высокая плодовитость и сложные жизненные циклы развития многих паразитов позволяют им выжить в борьбе за существование. Примером могут служить печеночный сосальщик (Fasciola hepatica), отличающийся сложным циклом развития и огромной плодовитостью; аскарида (Ascaris lumbriocoides), npoдуцирующая за 5–6 месяцев 50-60 млн. яиц, общий вес которых в 1700 раз превышает вес самки; свиной цепень (Taenia solium), выделяющий за год около 600 млн. яиц и живущий до 18 лет. Огромное количество семян развивается из цветка раффлезии, который достигает гигантских размеров – диаметр его около метра, а вес до 5 кг. Разумеется, из такого огромного количества яиц и семян какая-то часть паразитических организмов всегда найдет подходящие условия для развития.