

Глава 4. Совместное действие экологических факторов
Ограничивающий фактор. Все факторы в природе воздействуют на организм одновременно. Причем не в виде простой суммы, а как сложное взаимодействующее соотношение. Такая совокупность факторов называется их констелляцией. Поэтому оптимум и границы выносливости организма по отношению к какому-то одному фактору зависят от других воздействий. Например, при оптимальной температуре возрастает выносливость к неблагоприятной влажности, недостатку питания. С другой стороны, обилие пищи увеличивает устойчивость организма к изменениям нескольких климатических факторов. Однако эта так называемая «компенсация» факторов ограничена, и ни один из них не может быть полностью заменен другим. Вот почему при изменении того или иного условия жизнедеятельность организма (способность к конкуренции с другими видами, размножение и т. д.) лимитируется тем фактором, который сильнее отклоняется от оптимальной для вида величины. Если в количественном выражении хотя бы один из факторов выходит за пределы выносливости вида, то существование последнего становится невозможным, как бы ни были благоприятны остальные условия. Фактор, уровень которого в качественном или количественном отношении (недостаток или избыток) оказывается близким к пределам выносливости данного организма, называется ограничивающим.
Рассмотрим в качестве ограничивающего фактора температуру. Лось в Скандинавии встречается значительно севернее, чем в Сибири, хотя в последней средняя годовая температура выше. Причиной, препятствующей лосю расширить свой ареал на север в Сибири, оказываются низкие зимние температуры. Лимитирующим фактором распространения бука в Европе также является низкая температура января. Поэтому северные границы его ареала соответствуют январской изотерме -2°С. Рифообразующие кораллы обитают только в тропиках при температуре воды не ниже 20 °С.
Аналогичным фактором может быть и высокая температура. Так, южная граница ареала бабочки капустницы, широко распространенной в Европе и Северо-Западной Африке, находится в Палестине, поскольку летом там обычно слишком жарко.
При изменениях экологической обстановки нарушается и соотношение отдельных факторов. Вот почему в разных местностях факторы, ограничивающие развитие организмов, часто неодинаковы: на севере для определенных видов таковым может быть недостаток тепла, а на юге для тех же видов – недостаток влаги, пищи, высокая температура. Следует также отметить, что один и тот же фактор для одного организма некоторое время выступает как ограничивающий, а затем становится неограничивающим. Это зависит от стадии развития данного организма. Почти все животные и растения в период размножения более чувствительны к неблагоприятным условиям. Например, влияние климатических факторов при географическом расселении многих охотничье-промысловых птиц распространяется лишь на яйца и птенцов, но не на взрослых особей.
Экологические ряды и экологическая индивидуальность. Экологическим рядом называется совокупность растительных сообществ (фитоценозов), располагающихся соответственно нарастанию или убыванию какого-либо фактора (или группы факторов) среды. Например, на склоне наибольшая сухость почвы наблюдается в верхней части, а наименьшая – в нижней, поэтому здесь отмечаются различия в растительности, связанные с влажностью почвы. Одни виды произрастают только в верхней части склона, другие в средней, третьи в нижней. В результате четко выделяется экологический ряд видов растений либо по возрастанию либо по убыванию влажности почвы – сверху вниз от более к менее сухолюбивым и, наоборот, снизу вверх, от более к менее влаголюбивым. А экологический ряд древесных пород по возрастающей теневыносливости выглядит следующим образом: лиственница – береза – сосна – осина – ива – ольха серая – липа – дуб – ясень – клен – ольха черпая – ильм – граб – ель – бук – пихта.
Аналогичные экологические ряды составляются и по отношению растений к тепловому режиму, к степени засоленности почв, устойчивости к ветру и к другим факторам. Так, в поймах рек южной части Русской равнины в случае возвышения местности наблюдается смена растительности (от понижения на бугор) в такой последовательности: лугово-болотные, луговые, лугово-степные и степные растительные ассоциации. Это экологический ряд фитоценозов. Иногда в таком ряду выделяют до 10 и более ассоциаций. Границы их нередко очень трудно определить, поскольку сочетания экологических условий изменяются в пространстве постепенно и между ценозами образуется переходная, промежуточная, полоса, в которой совмещаются признаки соседствующих ассоциаций. Объясняется это экологической индивидуальностью каждого из видов, в связи с чем их ареалы в сообществе не совпадают. Иначе говоря, различные виды по-разному реагируют на одни и тс же факторы.
Вообще же экологическая индивидуальность особи - это совокупность специфических черт ее, заключающихся в своеобразном сочетании наследственных и приобретенных свойств. Она складывается в процессе развития организма (онтогенеза) и выражается в особенностях генотипа и фенотипа данной особи. В природе не существует одинаковых, тождественных, особей даже в очень однородной популяции. Кроме специфических признаков, каждая особь обладает и экологической индивидуальностью, проявляющейся в самых различных формах.
Среди большого количества особей, слагающих популяцию, всегда можно выделить индивидуумы наиболее или наименее экологически пластичные по отношению к тому или иному фактору. Одни очень чувствительны к понижению температуры, другие сравнительно выносливы к холоду, некоторые не выдерживают даже незначительной сухости, а есть и такие, которые выживают в засушливый период. Благодаря экологической индивидуальности в популяции обычно находятся самые жизнестойкие особи, переживающие весьма неблагоприятные условия, что обусловливает сохранение вида.
Правило предварения. В 1951 г. В. В. Алехин установил правило предварения для растений. Согласно этому правилу, северные влаголюбивые растения в пределах южных границ ареала располагаются на северных склонах и на дне балок, а южные по мере продвижения на север переходят на лучше прогреваемые южные склоны (рис. 4). Это особенно проявляется на южных и северных границах лесной зоны. По южным склонам из средней тайги глубоко в северную проникают ельники-черничники и ельники-кисличники. В Якутии на северных склонах растут холодовыносливые леса даурской лиственницы (Larix dahurica), а южные покрыты сосновыми лесами. На южных окраинах лесной зоны по северным склонам сохраняются леса, а на южных произрастает уже типичная степная растительность.
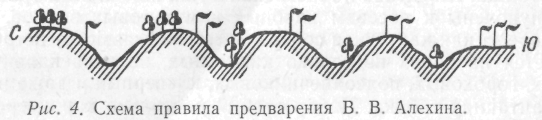
Естественно, что правило предварения носит относительный характер. Менее четко оно выражено в гористой местности, так как там отмечается более сложная совокупность экологических факторов. Тем не менее оно имеет большое значение при проведении геоботанических исследований, поскольку позволяет предсказывать состав растительности еще не обследованных мест и прежний ее облик там, где она уничтожена.
Принцип стадиальной верности. Под стацией обычно понимают место обитания вида. В связи с тем, что виды и слагающие их популяции избирательно относятся к факторам среды, они заселяют строго определенные стации с соответствующими экологическими условиями. Участок территории, занятый популяцией вида и характеризующийся определенными экологическими условиями, называется стацией. Понятие «стация» применяется только по отношению к виду.
Каждый вид имеет свой набор стаций. Между крайними показателями избирательности вида к местообитаниям существует много переходов. Азиатская саранча, например, живет только на болотистых стациях, а итальянская саранча (Calliptamus italicus) более пластична и заселяет целинные степные участки, залежные земли, пастбища. Шведская и гессенская мухи, пшеничный трипе приурочены к посевам хлебных или луговых злаков, в то время как капустная совка (Baraihra brassicae) встречается на полях не только капустных, но и свекловичных, гороховых, подсолнечниковых, клеверных и даже на плантациях табака. Набор стаций настолько характерен для каждого вида, что может служить не менее существенным отличительным признаком его, чем морфологические и другие особенности. Это имеет практическое значение при определении вредных и полезных видов.
Свойство видов избирательно заселять те или иные стации обозначается как принцип стациальной верности. Данный принцип является важной экологической закономерностью.
Правила смены местообитаний и ярусов. Принцип стадиальной верности применим лишь в условиях ограниченного пространства и времени. Закономерное изменение видами своих местообитаний в широком диапазоне пространства и времени является правилом смены местообитаний. Это правило было установлено и сформулировано Г. Я. Бей-Биенко (1966).
В свою очередь М. С. Гиляров вывел правило смены ярусов, показав, что в разных зонах одни и те же виды занимают неодинаковые ярусы. Это характерно для трансзональных видов, т. е. для видов, широко распространенных и встречающихся во многих природных зонах.
В пространстве правило смены местообитания выражается в зональной и вертикальной смене стаций и в зональной смене ярусов, а во времени – сезонной и годичной сменой стаций.
Зональная смена стаций – это закономерно направленное изменение местообитаний при переходе вида из одной природной зоны в другую. Обычно при продвижении на север виды избирают сухие, хорошо прогреваемые солнцем открытые стации с разреженным растительным покровом. Распространяясь к югу, эти же виды заселяют более увлажненные и тенистые места с густой растительностью. Например, перелетная саранча (Locusta migratoria) в Центральной Европе поселяется на песчаных местах, а в Средней Азии и Казахстане - на сырых болотистых с густым травостоем. На влажных лугах муравьи-лазии (Lasius niger, L. flavus) проявляют себя как гигрофобы и поселяются на кочках. В более сухих ареалах, в степи, эти же муравьи выступают как гигрофилы и выбирают более увлажненные стации. Как указывает Бей-Биенко, зональная смена стаций служит экологическим следствием закона географической зональности и объясняется изменением теплового режима. Внешне одинаковые стации на севере и на юге резко различаются именно по тепловому режиму, поэтому при продвижении с юга на север виды выбирают местообитания, приближающиеся по количеству тепла к южным.
Вертикальная смена стаций аналогична зональной, но она характерна для горных условий. Например, серый кузнечик (Decticus verrucivorus) в лесах Кавказа заселяет гигрофитные и мезофитные стации, а в альпийском поясе становится ксерофилом.
Зональная смена ярусов заключается в том, что многие виды при продвижении на север перемещаются из более высокого растительного яруса в более низкий, а некоторые в сравнительно сухих зонах из наземных становятся почвенными обитателями. Так, короед лесной садовник (Blastophagus piniperda) в центральных районах и на севере обитает под корой стволов и крупных ветвей сосны, а на юго-востоке европейской части СССР уходит в почву и поселяется на корнях. Личинки жука-оленя (Lucanus cervus) в лесной зоне развиваются в гниющих стволах и пнях, а в степной – в гнилых корнях на глубине до 100 см.
Бей-Биенко считает, что зональная смена стаций и ярусов и вертикальная смена стаций ставят вид в двойственные и противоречивые условия. С одной стороны, вид предъявляет к среде определенные требования, вытекающие из его наследственных физиологических свойств; с другой – при успешном расселении он вынужден занять новые стации или даже сменить ярус. В результате изменяется его экология, а вместе с тем и физиология. Следовательно, смена стаций становится одним из ведущих факторов эволюции.
Сезонная смена стаций происходит при колебаниях микроклимата в течение одного сезона. Наиболее четко это выражено в сухом и жарком климате и проявляется в переселении степных и пустынных видов в период засухи на посевы культурных растений, на луга, под полог леса, где сохраняются сравнительно высокая влажность и зеленый растительный покров. Такие миграции характерны для многих насекомых и грызунов.
Годичная смена стаций наблюдается при отклонении погодных условий от средней годовой нормы. Например, перелетная саранча в Южном Казахстане в сухие годы концентрируется на западинах с более влажной почвой и густым травяным покровом, а во влажные – заселяет возвышенные места.
Таким образом, смена местообитаний позволяет видам сохранять свой экологический стандарт в постоянно меняющихся условиях.
Принцип стадиальной верности и его противоположности – правила смены местообитаний и ярусов – свидетельствует о сложности взаимоотношений организмов со средой. Выяснение сущности этих взаимоотношений дает возможность более глубоко проникать в экологию того или иного вида и разрабатывать рациональные приемы по борьбе с вредными организмами и по охране и привлечению полезных.
Принципы экологической классификации организмов. Экологическая классификация организмов отличается от систематики тем, что в последней главным критерием служит филогенетическая близость организмов, т. е. систематика на всех ступенях таксономии базируется на едином критерии – филогенезе. В экологической классификации такого критерия нет, поэтому она имеет очень много схем.
Экологическая классификация организмов может быть проведена в соответствии с их положением в энергетической или пищевой цепи. По отношению к органическому веществу выделяются гетеротрофы и автотрофы, по функции в биогеоценозе – продуценты, консументы и редуценты (деструкторы).
В основу экологической классификации могут быть положены и места обитания.
Водные организмы при этом подразделяются на бентосные, планктонные и нектонные. Их можно классифицировать и по занимаемым зонам. При таком подходе важно выяснить положение организма во всех трех системах классификации, а также иметь в виду, что многие виды на разных стадиях развития ведут различный образ жизни (головастик и лягушка, личинка стрекозы и взрослое насекомое).
Особые затруднения вызывает классификация наземных животных, поскольку они представляют огромное разнообразие форм, что связано с особенностями мест обитания. Уже среди травоядных имеются и мелкие и очень крупные. Практически не поддается учету и экологической классификации обилие насекомых и других членистоногих, а также птиц. Еще сложнее классифицировать редуценты. Почвенные организмы обычно классифицируют по размерам, в связи с чем различаются микро-, мезо- и макробиота.
Наиболее распространена экологическая классификация организмов по жизненным формам, т. е. по типу внешней морфологии, отражающей важнейшие моменты образа жизни, отношение вида к среде. Жизненные формы определяют приспособленность организмов к комплексу факторов (в отличие от экологических групп, характеризующих приспособление к отдельным факторам), к специфике местообитания.
Очень многообразны жизненные формы у животных. Прежде всего это группы, обладающие похожими эколого-морфологическими приспособлениями для обитания в сходной среде. В данном случае термин «жизненные формы» заимствован из ботаники. Он утвердился в зоологии только в настоящем столетии, хотя животных издавна подразделяли на нырцов, норников, землероев и т. д.
Существует много различных трактовок жизненных форм животных. Это связано с тем, что за основу классификации в одних случаях берутся особенности размножения, в других – способы передвижения или добывания пищи. Нередко классификация базируется на приуроченности организмов к определенным экологическим нишам, ландшафту, ярусу. Тем не менее анализ жизненных форм дает возможность судить об особенностях среды обитания и путях выработки у животных адаптации к определенным условиям. Например, Д. Н. Кашкаров (1945) классифицирует жизненные формы животных следующим образом.
I. Плавающие формы:
Чисто водные: а) нектон, б) планктон, в) бентос;
Полуводные: а) ныряющие, б) неныряющие, в) добывающие из воды лишь пищу.
II. Роющие формы:
Абсолютные землерои (всю жизнь проводят под землей);
Относительные землерои (выходят на поверхность земли).
III. Наземные формы:
Не делающие нор: а) бегающие, б) прыгающие, в) ползающие;
Делающие норы: а) бегающие, б) прыгающие, в) ползающие;
Животные скал.
IV. Древесные, лазающие формы: а) не сходящие с деревьев, б) лишь лазающие по деревьям.
V. Воздушные формы: а) добывающие пищу в воздухе, б) высматривающие ее с воздуха.
Как видно, в основу этой классификации положены приспособления для передвижения. По отношению к влажности воздуха Кашкаров выделяет влаголюбивые (гигрофильные) и сухолюбивые (ксерофильные) формы; по питанию – растительноядных, всеядных, хищных, могильщиков (трупоедов); по месту размножения – размножающихся под землей, на поверхности земли, в ярусе трав, в кустарниках, на деревьях.
Различные категории жизненных форм насекомых относительно среды обитания (геобионты, гидробионты и т. д.) предлагает В. В. Яхонтов. Зонально-ландшафтную категорию жизненных форм разработали орнитологи А. К. Рустамов, Г. П. Дементьев, С. М. Успенский.
Растения классифицируют на основании адаптации к окружающим условиям. Среди них выделяются гигрофиты, мезофиты, ксерофиты. Данная классификация базируется на физиологических свойствах растений, а членение растительности па деревья, кустарники, траву дает характеристику главных наземных сообществ. В связи с многообразием условий на Земле у растений выработалось огромное количество жизненных форм. Понятие о жизненных формах растений впервые ввел в 1806 г. Гумбольдт. Обычно выделяют древесные, полудревесные, наземные травянистые и водные травянистые растения. Каждая из этих форм может быть представлена более мелкими группами. Самое широкое распространение получила классификация жизненых форм растений, разработанная в 1905–1907 гг. датским ботаником С. Раункиером. За ее основу принято расположение почек возобновления и наличие приспособлений для переживания неблагоприятного времени года. На этой классификации базируется современная классификация, в которой различают 6 жизненных форм растений (рис. 5).
Эпифиты* – воздушные растения, не имеющие корней в почве. Поселяются они на стволах других более крупных растений. В лесах это наствольные лишайники, реже мхи. Из высших растений эпифиты многочисленны во влажных тропических лесах.
Фанерофиты – надземные растения (деревья, кустарники, лианы, стеблевые суккуленты, травянисто-стеблевые растения). Почки возобновления находятся у них на вертикально расположенных побегах высоко над землей.
3. Хамефиты – травянистые растения с почками возобновления, расположенными у земли. В умеренных широтах побеги этих растений на зиму уходят под снег и не отмирают.

Гемикриптофиты – дернообразующие растения, у которых почки возобновления находятся на уровне почвы или даже в ней. Надземные побеги к зиме отмирают. Это очень многие луговые растения.
Криптофиты, или геофиты,– многолетние травы с отмирающими надземными частями. Почки возобновления располагаются на подземных органах (клубневые или корневищные растения).
Терофиты – однолетние растения. К зиме отмирают и надземные и подземные части их. Неблагоприятный период (зиму) переживают на стадии семян.
В приведенном ряду жизненных форм отчетливо проявляется возрастающая адаптация к неблагоприятным условиям. Во влажных тропических лесах большинство видов принадлежит к фанерофитам и эпифитам. В более северных районах преобладают растения с защищенными почками возобновления.
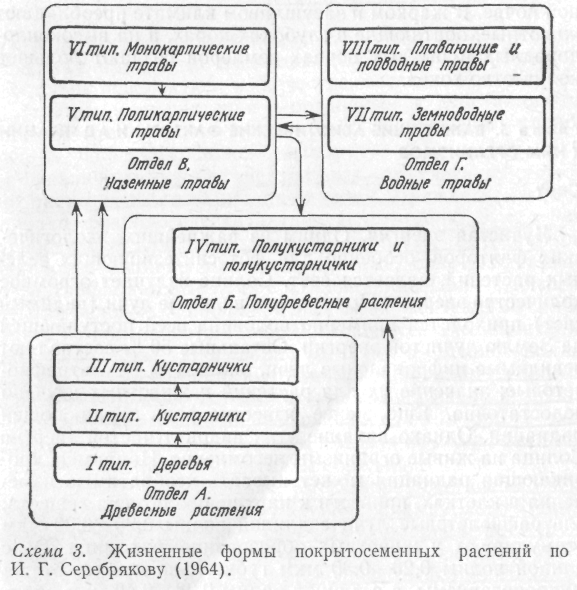
Существуют и другие схемы классификаций жизненных форм. Наибольшее признание получила классификация злаков по способу кущения, разработанная В. Р. Вильямсом. Г. Н. Высоцкий и Л. И. Казакевич в основу классификации жизненных форм положили характер подземных органов и способность растений к вегетативному размножению. В последнее время удачную классификацию покрытосеменных растений предложил И. Г. Серебряков, сосредоточив внимание на структуре и длительности жизни надземных скелетных осей. Он выделяет 4 отдела и 8 типов жизненных форм этих растений (схема 3). Каждый тип в свою очередь подразделяется на формы. К примеру, в I типе выделяются надземные кронообразующие деревья с прямостоячими стволами; кустовидные, одноствольные с низкими стволами; стланцы (с лежачими стволами).
Жизненные формы, доминирующие в том или ином сообществе, могут служить индикаторами условий обитания. Так, преобладание столонообразующих растений в широколиственных и темнохвойных лесах свидетельствует о малоплодородной, рыхлой и избыточно увлажненной почве. В жарком и засушливом климате преобладают животные, обитающие в глубоких норах, а на высокоплодородных и рыхлых почвах землерои создают большое количество ходов.