

Глава 13. Динамика и стабильность экосистем
Суточная и годовая динамика экосистем. Одной из характерных особенностей биоценозов является их суточная и годовая динамика. В каждом естественном биоценозе имеются группы организмов, активность жизни которых выпадает на разное время суток. Одни активны в ночное время и днем обычно скрываются в каких-либо убежищах. Другие, наоборот, пассивны ночью. В результате в составе и в соотношении отдельных видов биоценоза происходят периодические изменения, поскольку отдельные организмы на определенное время выключаются из него. При этом суточную динамику биоценоза обеспечивает не только животное, но и растительное население. У растений в течение суток также изменяются интенсивность и характер физиологических процессов, в ночные часы не происходит фотосинтез, у ряда растений цветки раскрываются только ночью и опыляются ночными животными, другие же приспособлены к опылению днем.
Большое значение в динамике биоценозов имеют суточные миграции животных. Они зависят от изменения характера действия различных факторов в течение суток. Особенно типичны суточные миграции морского планктона. Представители зоо- и фитопланктона в Каспийском море, например, днем держатся на глубине от 100 до 350 м, а ночью поднимаются в поверхностные слои. Вертикальные суточные миграции свойственны и почвенным обитателям.
Суточная динамика биогеоценозов в основном связана с ритмикой природных явлений и носит строго периодический характер.
Но, кроме этого, могут происходить и непериодические изменения активности и численности тех или иных компонентов экосистемы в течение суток, связанные с действием нерегулярных факторов среды. Так, сильные дожди или засухи приводят к перемещению животных, изменению их активности, влияют на интенсивность некоторых жизненных процессов у растений. Во время сильных ливней даже морской планктон из-за опреснения поверхностных слоев воды мигрирует в глубину.
Более существенные отклонения в биоценозах наблюдаются при сезонной динамике. Они обусловлены биологическими циклами организмов, зависящими от сезонной цикличности природных явлений. Смена времен года оказывает значительное влияние на жизнедеятельность растений и животных (периоды цветения, плодоношения, активного роста, листопада и зимнего покоя у растений; спячка, зимний сон, диапауза и миграции у животных).
В разных широтах длительность биологических сезонов неодинакова. Поэтому сезонная динамика биогеоценозов арктической, умеренной и тропической зон различна. Наиболее четко она выражена в экосистемах умеренного климата и в северных широтах.
В связи с тем что характер суточных и сезонных изменений более или менее постоянен в течение длительного времени (столетия и даже тысячелетия), в биоценозах исторически сформировались механизмы, приводящие сообщество в целом в соответствие с периодикой изменения условий обитания.
Экологические сукцессии. В процессе суточной и сезонной динамики целостность биоценозов обычно не нарушается. Биоценоз испытывает лишь периодические колебания качественных и количественных характеристик. Но он может подвергаться и воздействию сил, которые существенно изменяют или полностью разрушают сообщество. В таких случаях развивается другой, более приспособленный к новым условиям биоценоз. Подобная последовательная смена одного биоценоза другим называется экологической сукцессией (succession – последовательность, смена). В целом цепь сменяющих друг друга биоценозов называется сукцессионным рядом или серией (сериальные стадии). В сукцессионном ряду каждый биоценоз представляет собой определенную стадию формирования конечного, завершающего, или так называемого климаксного сообщества. По мере усложнения сообщества усложняются и связи между популяциями. Менее приспособленные к новым условиям замещаются более приспособленными, и так до тех пор, пока не появятся виды, которых условия среды полностью устраивают и они уже не замещаются другими видами. В результате сообщество становится стабильным и достигает своей завершающей стадии.
Концепция о заключительной, завершающей фазе развития биоценоза, точнее биогеоценоза, как учение о климаксе имеет некоторые негативные стороны. Дело в том, что в любом биогеоценозе действуют определенные противоречивые движущие силы развития. Различают противоречия между синтезирующими и разрушающими органические вещества группами организмов (внутренние противоречия) и противоречия между биоценозом и опосредствованной им средой обитания (внешние противоречия). Единство этих двух типов противоречий лежит в основе существования и развития биоценозов на Земле. Поэтому сами биоценозы также относительно устойчивы. Выражается эта относительность и противоречивость в колебаниях численности отдельных популяций, особенно в периоды вспышек массового размножения, во время вторжения новых, устойчивых видов и т. д. В результате могут нарушаться биоценотические связи и происходить заметные, иногда необратимые, изменения в строении биоценозов.
С позиций диалектического материализма существование абсолютно климаксного биоценоза невозможно, так как взаимное воздействие компонентов его никогда не прекращается, а следовательно, и не останавливается биогеоценотический процесс. В результате совершаются сукцессии. Этот процесс постоянен, ибо развитие биоценозов, как и любого природного явления, не может остановиться. Со временем микроорганизмы, растения и животные в биогеоценозе (его составная живая часть – биоценоз) приобретают некоторую относительную устойчивость – динамическое равновесие, сбалансированность отдельных компонентов в сообществе. По такое климаксное состояние биоценоза не бывает постоянным во времени и пространстве, раз и навсегда установившимся. Поэтому термин «климакс» можно использовать лишь условно в смысле приобретения биоценозом определенной замедленности в развитии, поскольку даже при кажущейся неизменности в биоценозе все-таки протекают процессы, подготавливающие его к сукцессии.
В историческом разрезе смена фауны и флоры по геологическом периодам не что иное, как смена сообществ, замена одного типа биоценоза другим, т. е. – экологическая сукцессия. Эти сукцессии тесно связаны с геологическим и климатическими изменениями, а также с эволюцией видов.
Нередко сукцессии совершаются в сравнительно короткие промежутки времени и тогда они не связаны с эволюцией видов. Ведь биоценоз в течение ряда лет не остается постоянным. Изменяются условия жизни, исчезают одни и появляются другие группы организмов, меняются связи между популяциями, а вслед за этим – и биоценоз в целом. Иногда такие изменения происходят очень медленно, даже столетиями. А бывает, что и относительно быстро. Лесной пожар может моментально уничтожить сложившийся на протяжении тысячелетий устойчивый биоценоз, и на пожарище начинает довольно быстро создаваться новое сообщество. При этом будет наблюдаться частая смена ряда сообществ. В конечном итоге устойчиво восстанавливается биоценоз исходного типа (или же он незначительно отличается от первоначального). Классическим примером сукцессии с образованием устойчивого биоценоза является зарастание озера и возникновение на его месте торфяного болота или формирование елового леса на брошенных землях (рис.43).
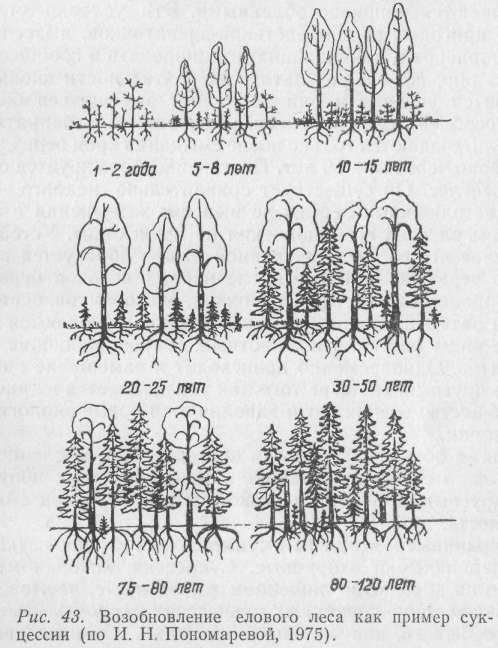
Еловый лес в своем развитии проходит несколько этапов. Первыми на заброшенной пашне из древесных пород появляются береза, осина и ольха, поскольку семена этих деревьев легко разносятся ветром. Попав на слабо задернованную почву, они прорастают. Такие первопоселенцы обычно называются пионерами. Наиболее стойкие из них заселяют заброшенную или распаханную территорию, утверждаются там и постепенно изменяют среду, создавая новые условия, к которым сами со временем оказываются неприспособленными. Эти условия становятся пригодными для растений-захватчиков, вытесняющих пионеров и начинающих доминировать в сообществе до тех пор, пока в результате их деятельности вновь не изменятся условия и они не начнут замещаться более приспособленными формами. Условия, благоприятные для ели, создаются только после смыкания крон берез, т. е. примерно через 30–50 лет. Постепенно формируется смешанный лес. Он существует сравнительно недолго, так как светолюбивые березы не выносят затемнения и под пологом елей их возобновления не происходит. Устойчивый еловый лес на заброшенной пашне образуется примерно через 80–120 лет после первых всходов березы.
В процессе развития березняков, ольховников, осинников, а затем и елового леса в биоценоз включаются все новые виды растений и животных, сопровождающие доминантов. Одновременно происходит и замещение одних видов другими. По мере того как увеличивается их число, в сообществе возникают и заполняются новые экологические ниши.
Таким образом, сукцессия протекает как медленное и в какой-то степени случайное замещение одних популяций другими, а не путем резкой, скачкообразной смены сообществ.
Первичные и вторичные сукцессии. Различают сукцессии первичные и вторичные. Сукцессия, которая начинается на абсолютно лишенном жизни месте, называется первичной. При первичных сукцессиях скорость изменения сообществ, как правило, невелика. Сукцессионные ряды, последовательно сменяющие друг друга, удерживаются значительный промежуток времени, а достижение биоценозом климаксного состояния затягивается иногда на столетия и больше. К типичным первичным сукцессионным образованиям биоценозов в масштабе геологических периодов относится формирование фауны и флоры на вновь появившейся песчаной дюне, на лавовом потоке, в послеледниковый период, после поднятия материков и т. д.
Если сообщество развивается на месте, где ранее существовал хорошо развитый биоценоз, то сукцессия будет вторичной. В таких местах обычно сохраняются богатые жизненные ресурсы. Поэтому вторичные сукцессии приводят к образованию климаксного сообщества значительно быстрей, чем первичные. В современных условиях вторичные сукцессии наблюдаются повсеместно. Они обусловлены последствиями, возникающими в результате пожаров, наводнений, распашки степей, вырубки лесов, осушения болот.
Протекают сукцессии и внутри биоценозов. Примером быстрых сукцессии комплексов организмов местного характера на мелких участках являются дупла деревьев. Дупла обладают сравнительно устойчивым микроклиматом. От величины дупла, толщины его стенок, состояния древесины зависят условия, благоприятствующие заселению его теми или иными видами организмов. Постепенно, в процессе сукцессии, в дупле формируется своеобразный биоценоз. При заселении дупла каким-либо видом птиц или млекопитающих сообщество мелких организмов претерпевает коренную перестройку. Вокруг основного обитателя дупла концентрируются сопутствующие виды (квартиранты, паразиты, симбионты, комменсалы). Между хозяином дупла и остальными жителями его устанавливаются строго определенные количественные пропорции. В результате на какой-то период образуется своеобразное климаксное сообщество.
Еще более характерные сукцессии протекают в норах. Лишенная обитателей нора имеет температуру и влажность, близкие к окружающей среде. Если же нора обитаема, то в ней создается вполне специфический микроклимат. Норовые биоценозы нередко насчитывают до 200 и более квартирантов. Многие из них связаны с хозяином норы не только пространственными, но и пищевыми взаимоотношениями (паразиты). Естественно, что устойчивый норовый биоценоз развивается постепенно, в процессе сменяющих друг друга сукцессионных рядов.
Хорошо изучены сукцессии в разлагающихся стволах деревьев. В течение первых 3–4 лет древесину заселяют в основном ксилофаги из насекомых и деревоокрашивающие грибы. Позже появляются дереворазрушающие грибы, увеличивается количество ксилофагов, хищных насекомых. Затем в гниющей древесине поселяются сапроксилофаги, т. е. насекомые, питающиеся гнилой древесиной. Вместе с этим растет численность хищников и изменяется их видовой состав. На 8–10-м году количество ксилофагов начинает сокращаться, соответственно снижается и число хищников, а число сапроксилофагов возрастает. Затем появляются шляпочные грибы. Примерно через 10–15 лет древесина полностью разрушается и специфичный биоценоз исчезает.
Интересна динамика биоценозов в слое растительного опада, при обрастании камней, заселении трупов животных, высыхающего навоза. Например, трупы животных на различных стадиях разложения заселяются разными комплексами организмов. Из насекомых вначале появляются могильщики и мертвоеды (Necrophorus, Silpha), затем мухи (Calliphoridae, Muscidae, Sarcophagidae) и жуки-карапузики (Histeridae). Уже высохшие трупы заселяются кожеедами (Dermestidae).
Известны сукцессии биоценозов плодовых деревьев по мере изменения их состояния. Молодые неплодоносящие сады заселяются многоядными насекомыми-фитофагами, среди которых наиболее характерны хрущи (Melolonthinае) и чернотелки (Tenebrionidae); из бабочек – зимняя пяденица (Operoptera brumata), кольчатый шелкопряд (Malacosoma neustria), различные листовертки (Tortricidae). Кроме этого, многочисленны тли (Aphidodea) и щитовки (Diaspidae). Когда сад начинает плодоносить, появляются насекомые, питающиеся цветками и плодами (яблонная и другие плодожорки – Laspeyresia, Grapholitha, яблонный цветоед – Anthonomus pomorum, вишневый слоник – Rhynchites auratus, казарка – R. bacchus). По мере старения сада в нем обнаруживаются стволовые вредители (короеды – Ipidae, древоточцы – Cossidae, стеклянницы – Aegeriidae). Численность их постоянно растет. Вслед за изменением фитофагов меняется в количественном и качественном отношении и фауна паразитов и хищников.
Общие закономерности сукцессии. Экологическая сукцессия является результатом изменений, которые вносятся в среду обитания самими сообществами. Это закономерный, направленный процесс. Он заканчивается образованием климаксного биоценоза, характеризующегося максимальной величиной биомассы и наибольшим разнообразием слагающих его видов. Климаксный биоценоз находится в состоянии гомеостаза.
Общие закономерности сукцессии, согласно Ю. Одуму, сводятся к следующему. В направлении сукцессии от первичных стадий к климаксу, или от юных к стареющим сообществам, изменяется видовая и органическая структура, а также миграция энергии – метаболизм сообщества.
Видовой состав в сукцессионном ряду меняется сначала быстро, а затем более медленно. Видовая численность автотрофов увеличивается в первичных и часто в начале вторичных сукцессии, хотя на стадиях старения может снижаться. Количество видов гетеротрофов растет до относительно поздней стадии. Таким образом, видовое разнообразие поначалу увеличивается, потом стабилизируется и в итоге, на стадиях старения, снижается.
При изменении органической структуры сукцессии увеличивается общая биомасса сообщества и количество мертвого органического вещества, в течение ранней фазы первичной сукцессии возрастает количество хлорофилла.
Метаболизм (миграция энергии) по ходу смены сообществ также изменяется. Связи по линии пищевой цепи становятся все более сложными. Общая продуктивность повышается в течение ранней фазы первичной сукцессии и мало или совсем не изменяется во время вторичной сукцессии. Чистая продуктивность сообщества снижается, а дыхание его растет.
Ю. Одум указывает, что наибольшее значение в перечисленных закономерностях имеют 4 аспекта.
Видовой состав растений и животных в процессе сукцессии непрерывно меняется. Это свидетельствует о том, что виды, доминирующие на начальных стадиях сукцессии, по мере достижения биоценозом климаксного состояния постепенно утрачивают это свойство. Такое явление наглядно прослеживается на рассмотренном выше примере с заменой березняка еловым лесом.
Биомасса органического вещества увеличивается по ходу сукцессии. Совершенно ясно, что с течением времени как в наземной, так и в водной среде, происходит накопление разлагающегося органического вещества. Естественные водоемы часто содержат в растворенном состоянии столько же органического вещества, сколько его находится в теле живых организмов.
Видовое разнообразие имеет тенденцию увеличиваться по ходу сукцессии. Это тесно связано со вторым аспектом, поскольку по мере роста биомассы растет число экологических ниш (лесная подстилка, гумус, кора, сухостой). В результате наиболее интенсивно увеличивается видовое разнообразие гетеротрофов. Автотрофы достигают этого на сравнительно ранних этапах сукцессии и обеспечивают максимум первичной продукции, создавая тем самым благоприятные условия для увеличения разнообразия гетеротрофов.
Снижение чистой продукции сообщества и соответственное повышение интенсивности дыхания – самые важные явления сукцессии. Происходит это по следующим причинам. Максимальная продуктивность растений наблюдается тогда, когда суммарная поверхность листьев, освещаемая солнечным светом, в 4–5 раз больше поверхности грунта. Увеличение массы листьев не влечет за собой увеличения фотосинтеза, так как повышенная затененность снижает полезную площадь фотосинтезирующей поверхности. Поэтому дыхание затененных листьев отрицательно влияет на чистую продуктивность фотосинтеза. В лесах листовая поверхность, отнесенная к поверхности почвы, примерно в 10 раз больше. Однако леса считаются преуспевающими биогеоценозами, а это позволяет предполагать, что большая масса листьев не только играет роль в процессах созидания первичной продукции, но и выполняет другие важные функции.
Таким образом, продуктивность сообщества на разных стадиях сукцессии различна. На начальных стадиях приход ее (созидание органического энергетического материала) превышает расход (затраты на дыхание). В результате быстро увеличивается биомасса сообщества. По мере приближения сукцессии к климаксному уровню продуктивность, достигнув максимума, начинает уменьшаться. В климаксном биоценозе продуктивность и дыхание (приход и расход) уравновешиваются и увеличения как первого, так и второго процессов не наблюдается: биоценоз достигает более или менее стабильного, гомеостатического, состояния. Это означает, что в климаксном биоценозе все создаваемые пищевые вещества используются, вследствие чего чистая продукция остается невысокой.
В стабильных климаксных биоценозах валовая первичная продукция обычно полностью расходуется на автотрофное и гетеротрофное дыхание и чистой продукции сообщества в конце года очень мало или ее совсем не остается. Достижение климакса, стабильности, проявляется в сбалансированности автотрофности и гетеротрофности. В идеальном случае автотрофность становится равной гетеротрофности.
Несколько иначе обстоит дело при так называемых гетеротрофных сукцессиях, протекающих внутри биоценоза (в разлагающихся стволах деревьев, на трупах животных и др.). В срубленном дереве, например, энергетические запасы вначале максимальны, но по мере сукцессии снижаются, поскольку автотрофный режим здесь исключен. В результате из-за отсутствия автотрофного процесса в таких биоценозах не наступает климаксного состояния и сообщество, достигнув какого-то максимума, через сравнительно короткий промежуток времени, после того как будут исчерпаны энергетические запасы, разрушается и исчезает.
Стабильные и нестабильные биоценозы. Важнейшей особенностью любого естественного биоценоза является его способность к саморегулированию, т. е. к удержанию основных параметров во времени и пространстве. Относительная стабильность биоценоза обеспечивает устойчивый круговорот веществ и поток энергии. В развитом биоценозе возникает некоторое равновесие между создаваемой и потребляемой продукцией.
Наиболее стабильны биоценозы, достигшие климаксного состояния. При этом стабильность биоценоза находится в прямой зависимости от его сложности. Чем больше видовое разнообразие биоценоза, тем он стабильнее. В таких биоценозах формируются сложные пищевые взаимоотношения, сложные циклы (сети) питания. Биоценозы с упрощенной структурой крайне неустойчивы, в них происходят резкие колебания численности отдельных популяций.
В биоценозах в случае резкого увеличения численности популяции какого-либо вида немедленно вступают в действие механизмы, регулирующие ее. Возможностей для такой регуляции в сложных биоценозах значительно больше, чем в простых, поскольку между видами имеется множество пищевых связей, одна форма которых в случае необходимости компенсируется другой. Подтверждений этому в природе очень много. Сложные биоценозы тропических лесов, например, исключительно стабильны, в то время как в Арктике недостаток видов, способных заменить в качестве пищи основной вид, приводит к резкому колебанию численности популяций. Тундровые упрощенные биоценозы в сравнении с тропическими, как правило, менее устойчивы.
В естественных условиях через любую биологическую систему, в том числе и биоценоз, постоянно проходит поток энергии. Согласно второму закону термодинамики, рассеивание энергии связано с принципом стабильности. В силу этого любой естественный биоценоз развивается в направлении устойчивого состояния. Это было показано на примере сукцессии, ведущих к климаксу. И совершенно ясно, что чем больше биомасса того или иного биоценоза, тем больше должны быть затраты на поддержание его стабильности. Однако если общая биомасса распадается на достаточно большое количество видов с разными свойствами, то затраты на поддержание стабильного состояния каждой структурной единицы (популяции, особь) сокращаются. Этим и объясняется тенденция любого естественного биоценоза в процессе сукцессии увеличивать свое видовое разнообразие.
Несколько по-иному проявляется стабильность во вторичных агробиоценозах, существенно отличающихся от естественных, первичных, не только ненормально высоким доминированием небольшого количества видов, но и тем, что численность этих видов поддерживается деятельностью человека, который в форме урожая постоянно и регулярно изымает из них биологическую продукцию, сохраняя тем самым относительную устойчивость агробиоценозов.
Как считает М. С. Гиляров, по структуре агробиоценозы приближаются к тем естественным биоценозам, которые формируются в экстремальных условиях. Отличаются они лишь тем, что их популяции (комплексы популяций) не развиваются до саморегулирующихся систем.
Степень устойчивости агробиоценозов зависит от частоты и характера воздействий человека.
Вместе с тем между первичными и вторичными биоценозами все же нет четких границ и можно проследить все переходы от пастбища, сенокоса, сеяной травосмеси до зерновых и пропашных культур. В этом проявляется и последовательность все усиливающегося антропического влияния.
В целом естественные биоценозы, как правило, более стабильны и обладают большим видовым разнообразием, чем агробиоценозы. Последним свойственна монотонность, тенденция к катастрофическим вспышкам массового размножения вредителей, к снижению урожайности.
Изучение сельскохозяйственных и лесных биоценозов показывает, что обеднение их видового состава приводит к тому, что они сильнее и чаще страдают от вспышек массового размножения вредителей. Это особенно характерно для монокультур.