

Часть IV
Экология сообществ и экосистем
Глава 9. Биотические связи организмов в биоценозах
Основные понятия синэкологии. В природе популяции разных видов интегрируются в макросистемы более высокого ранга – в так называемые сообщества, или биоценозы. Биоценоз (от гр. bios – жизнь, koinos – общий) – это организованная группа популяций растений, животных и микроорганизмов, живущих, совместно в одних и тех же условиях среды. Понятие «биоценоз» было предложено в 1877 г. немецким зоологом К. Мебиусом. Мебиус, изучая устричные банки, пришел к выводу, что каждая из них представляет собой сообщество живых существ, все члены которого находятся в тесной взаимосвязи. Биоценоз является продуктом естественного отбора. Выживание его, устойчивое существование во времени и пространстве зависит от характера взаимодействия составляющих популяций и возможно лишь при обязательном поступлении извне лучистой энергии Солнца.
Но никакой биоценоз не может развиваться сам по себе, вне и независимо от среды. В результате в природе складываются определенные комплексы, совокупности живых и неживых компонентов, где сложные взаимодействия отдельных частей их поддерживаются на основе разносторонней взаимной приспособленности. Пространство с более или менее однородными условиями, заселенное тем или иным сообществом организмов (биоценозом), называется биотопом. Иначе говоря, биотоп – это место существования, местообитание, биоценоза. Поэтому биоценоз можно рассматривать как исторически сложившийся комплекс организмов, характерный для какого-то конкретного биотопа.
Любой биоценоз образует с биотопом диалектическое единство, биологическую макросистему еще более высокого ранга – биогеоценоз. Термин «биогеоценоз» предложил в 1940 г. В. Н. Сукачев. Он практически тождествен широко распространенному за рубежом термину «экосистема», который был предложен в 1935 г. А. Тенсли. Существует мнение, будто содержание термина «биогеоценоз» в значительно большей степени отражает структурные характеристики изучаемой макросистемы, тогда как в понятие «экосистема» вкладывается прежде всего ее функциональная сущность. Фактически между этими терминами различий нет. Несомненно, В. Н. Сукачев, формулируя понятие «биогеоценоз», объединял в его содержании не только структурную, но и функциональную значимость макросистемы. По В. Н. Сукачеву, биогеоценоз – это совокупность на известном протяжении земной поверхности однородных природных явлений – атмосферы, горной породы, гидрологических условий, растительности, животного мира, мира микроорганизмов и почвы. Эта совокупность отличается спецификой взаимодействий слагающих ее компонентов, их особой структурой и определенным типом обмена веществ и энергии между собой и с другими явлениями природы.
Биогеоценозы могут быть самых различных размеров. Кроме того, они отличаются большой сложностью – в них подчас трудно учесть все элементы, все звенья. Это, к примеру, такие естественные группировки, как лес, озеро, луг и т. д. Примером сравнительно простого и четкого биогеоценоза может служить небольшой водоем, пруд. К неживым компонентам его относятся вода, растворенные в ней вещества (кислород, углекислый газ, соли, органические соединения) и грунт – дно водоема, где также содержится большое количество разнообразных веществ. Живые компоненты водоема разделяются на производителей первичной продукции – продуценты (зеленые растения), потребителей – консументы (первичные – растительноядные животные, вторичные – плотоядные животные и т. д.) и разрушителей – деструкторы (микроорганизмы), которые разлагают органические соединения до неорганических. Любой биогеоценоз, независимо от его размеров и сложности, состоит из этих основных звеньев: производителей, потребителей, разрушителей и компонентов неживой природы, а также из множества других звеньев. Между ними возникают связи самых различных порядков – параллельные и перекрещивающиеся, запутанные и переплетенные и т. д.
Сбалансированные животно-растительные сообщества (биоценозы) являются высшей формой существования организмов. Они характеризуются относительно устойчивым составом фауны и флоры и обладают типичным набором живых организмов, удерживающим свои основные признаки во времени и пространстве. Устойчивость биогеоценозов поддерживается саморегуляцией, т. е. все элементы системы существуют совместно, никогда полностью не уничтожая друг друга, а только ограничивая численность особей каждого вида до какого-то предела. Именно поэтому между видами животных, растений и микроорганизмов исторически сложились такие взаимоотношения, которые обеспечивают развитие и удерживают размножение их на определенном уровне. Перенаселенность одного из них может возникнуть по какой-то причине как вспышка массового размножения, и тогда сложившееся соотношение между видами временно нарушается.
В целом биогеоценоз представляет внутреннее противоречивое диалектическое единство, находящееся в постоянном движении и изменении. «Биогеоценоз – не сумма биоценоза и среды,– указывает Н. В. Дылис,– а целостное и качественно обособленное явление природы, действующее и развивающееся по своим собственным закономерностям, основу которых составляет метаболизм его компонентов».
Чтобы упростить изучение биоценоза, его условно можно расчленить на отдельные компоненты: фитоценоз – растительность, зооценоз – животный мир, микробиоценоз – микроорганизмы. Но такое дробление приводит к искусственному и фактически неправильному выделению из единого природного комплекса группировок, которые самостоятельно существовать не могут. Ни в одном местообитании не может быть динамической системы, которая состояла бы только из растений или только из животных. Биоценоз, фитоценоз и зооценоз необходимо рассматривать как биологические единства разных типов и степеней. Такой взгляд объективно отражает реальное положение в современной биологии.
В. А. Догель, Ε. Η. Павловский, представляя организм как среду жизни паразитов, разработали стройное учение о паразитоценозе. Организм хозяина, согласно этому учению, служит биотопом, местообитанием паразитоценоза – своеобразной группировки организмов, вступающей со своим биотопом в специфические взаимоотношения. Но так как хозяин всегда обитает в сложном многовидовом сообществе, паразитоценоз может рассматриваться как компонент биоценоза.
В современных условиях деятельность человека преобразует природные биогеоценозы (леса, степи, озера). На смену им приходят посевы и посадки культурных растений. Так формируются особые вторичные агробиогеоценозы, или агроценозы, количество которых на Земле постоянно увеличивается. Агроценозами являются не только сельскохозяйственные поля, но и полезащитные лесные полосы, пастбища, искуственно возобновляемые леса на вырубках и пожарищах, пруды и водохранилища, каналы и осушенные болота. Агробиоценозы по своей структуре характеризуются незначительным количеством видов, но высокой их численностью. Хотя в структуре и энергетике естественных и искусственных биоценозов есть много специфичных черт, резких различий между ними не существует. В естественном биогеоценозе количественное соотношение особей разных видов взаимно обусловлено, поскольку в нем действуют механизмы, регулирующие это соотношение. В результате в таких биогеоценозах устанавливается динамическое равновесие, поддерживающее наиболее выгодные количественные пропорции составляющих его компонентов. В искусственных агроценозах нет подобных механизмов, там человек полностью взял на себя заботу об уравновешивании взаимоотношений между видами. Изучению структуры и динамики агроценозов уделяется большое внимание, так как уже в обозримом будущем первичных, естественных, биогеоценозов практически не останется.
Общая характеристика биотических факторов. Биотические факторы – это совокупность влияний жизнедеятельности одних организмов на другие. Эти факторы носят самый разнообразный характер и проявляются во взаимоотношениях организмов при совместном обитании. Так, растения представляют собой важный биотический фактор для животных-фитофагов – от их количества зависит численность и распространение последних. В свою очередь фитофаги оказывают влияние на растения - снижают их продуктивность и создают неблагоприятные условия для размножения.
Определенную среду обитания для многих животных создают растительные ассоциации. Хорошо известны типичные лесные или степные животные, приспособленные к жизни либо в густом лесу, либо в травянистой степной растительности.
Важным биотическим фактором для ряда цветковых растений выступают животные-опылители. Взаимные приспособления цветков и животных, опыляющих их, выработавшиеся в процессе длительного совместного существования, часто бывают прямо-таки изумительными. Не менее поразительны взаимные приспособления у плодов растений и животных, способствующих их расселению (зоохория).
Хищники регулируют динамику численности своих жертв. С другой стороны, то или иное количество добычи обусловливает динамику численности хищников.
Весьма интересные взаимоотношения складываются у паразитических видов со средой обитания, которой служит организм хозяина. В данном случае и паразит и хозяин взаимно приспосабливаются к совместному существованию.
Взаимоотношения организмов выражаются также и в химическом воздействии их друг на друга. Выделяемые растениями колины, фитонциды и другие биологически активные вещества оказывают либо угнетающее, либо стимулирующее действие на различные организмы. Выделяемые микроорганизмами антибиотики также меняют условия жизни многих организмов. Кроме того, организмы могут механически воздействовать друг на друга. Например, вытаптывание животными травяного покрова.
Действие биотических факторов может быть и косвенным. Живые организмы в процессе жизнедеятельности вносят изменения в окружающую неживую природу, а это влечет за собой смену условий среды. Так, бактерии, влияя на состав почвы, изменяют условия существования растений и почвенных животных. Под пологом леса, на лугу и в других местообитаниях формируются специфичные микроклиматы, где одни организмы находят благоприятные факторы, а другие нет.
Типы биотических отношений. В биоценозах между различными видами организмов возникают определенные связи. Основной формой этих связей служат пищевые взаимоотношения, на базе которых формируются сложные цепи и циклы питания. Кроме пищевых, в сообществе возникают пространственные связи. И именно на пищевых и пространственных отношениях строятся разнообразные биотические комплексы, объединяющие виды в единое целое, в биологическую макросистему. Основные формы биотических отношений могут быть сведены к следующим.
Конкуренция – межвидовые взаимоотношения, при которых популяции в борьбе за пищу, местообитание и другие необходимые для жизни условия воздействуют друг на друга отрицательно. Классическим примером конкуренции являются описанные Г. Ф. Гаузе (см. гл. 10) опыты по содержанию популяций двух видов инфузорий на одном и том же ограниченном питании. Как оказалось, через некоторое время в живых остаются особи только одного вида, выжившие в борьбе за пищу, поскольку его популяция быстрее росла и размножалась. В данном случае конкуренция обусловлена ограниченными пищевыми ресурсами. Выведенное из этих исследований правило, так называемая теорема Гаузе, заключается в том, что два одинаковых в экологическом отношении вида сосуществовать не могут, что конкуренция особенно сурова между животными, обладающими сходными экологическими потребностями.
Обостренной конкуренцией нередко объясняется несовместимость в одном водоеме широкопалого (Astacus astacus) и узкопалого (A. leptodactylus) раков. Обычно победителем оказывается узкопалый рак как наиболее плодовитый и приспособленный к современным условиям жизни.
Примером конкуренции растений могут служить взаимоотношения лисохвоста лугового (Alopecurus ргаtensis) и типчака (Festuca sulcata). Эксперименты показали, что типчак может произрастать во влажной почве, но в сообществе лисохвостного луга не растет. Здесь он подавляется теневыносливым и быстро растущим лисохвостом. Таким образом, в формировании типчакового или лисохвостного фитоценоза решающее значение имеет не влажность почвы, а конкурентные взаимоотношения типчака и лисохвоста. В более сухих местообитаниях типчак заглушает лисохвост, а на увлажненных лугах лисохвост выходит победителем.
Конкуренция является одной из причин того, что два вида, слабо различающихся спецификой питания, поведения, образа жизни и т. д., редко сожительствуют в одном сообществе. Такая конкуренция носит характер прямой вражды.
Чаще же конкуренция проявляется косвенно. В биоценозах она незначительна, поскольку различные виды, занимая определенные экологические ниши, неодинаково воспринимают одни и те же факторы среды. Чем разнообразнее возможность организмов биоценоза, тем менее напряженной будет конкуренция.
Хищничество – форма межвидовых взаимоотношений, при которой один вид живет за счет другого, нанося ему ущерб. В основе этих отношений также лежат пищевые связи. Как правило, хищник вначале убивает добычу, а затем поедает ее. Но прежде он должен поймать жертву, и для этого у него есть специальные приспособления. Однако и у жертв исторически вырабатываются защитные свойства в виде анатомо-морфологических, физиологических, биохимических и других особенностей. Это выросты тела, шипы, колючки, панцири, защитная окраска, ядовитые железы, способность быстро прятаться, зарываться в рыхлый грунт, строить недоступные хищникам убежища, прибегать к сигнализации об опасностях. В результате таких обоюдных приспособлений формируются вполне определенные группировки организмов – специализированные хищники и специализированные жертвы. Так, коршун-слизнеед (Rosthramus sociabilis) употребляет в пищу только некоторые виды моллюсков, скопа (Pandion holiaetus) питается рыбой, основной пищей рыси (Felix lynx) служат зайцы, а волк (Canis lupus)–типичный многоядный хищник. Однако во всех этих отношениях в процессе взаимных приспособлений постоянно происходит эволюция и естественный отбор. И как результат исторического развития взаимоотношений хищник – жертва в любом биоценозе сформировались определенные механизмы регуляции численности обоих компонентов системы. Поэтому она всегда удерживается в пределах какой-то величины, приближающейся к оптимуму плотности популяций как хищника, так и его добычи.
До недавнего времени было широко распространено мнение, будто все хищники – вредные животные и их следует уничтожать. Это ошибочное представление, поскольку уничтожение хищников часто приводит к нежелательным последствиям и наносит большой ущерб и дикой природе и хозяйству человека. Ведь жертвами хищника обычно бывают больные и ослабленные особи, уничтожением которых сдерживается распространение болезней и эпизоотии, оздоравливается та или иная популяция. Сейчас уже нет сомнения, что волки, например, содействуют интенсивному размножению и повышению жизнеспособности популяции северных оленей в лесотундре и тундре; щуки в прудовых хозяйствах, если они количественно не превышают какого-то предела, стимулируют продуктивность карпов.
Иначе говоря, хищник – важный фактор естественного отбора. Под его прессом постоянно улучшается состав популяций тех или иных организмов, и он в какой-то степени обусловливает их прогрессивное развитие. С другой стороны, жертвы также принимают активное участие в этом процессе и оказывают влияние на своих хищников, содействуя их совершенствованию и прогрессу. Следовательно, эта борьба взаимно противоположных начал является движущей силой эволюции как хищника, так и жертвы.
Паразитизм – межвидовые взаимоотношения, при которых один вид живет за счет другого (хозяин), поселяясь внутри или на поверхности его тела. Паразитизм возник в процессе тесного контакта различных видов организмов также на базе пищевых и пространственных связей и характерен для многих организмов, но наиболее широко распространен среди низших и мелких растений и животных – вирусов, бактерий, грибов, простейших, червей, в несколько меньшей степени – среди членистоногих.
Паразитизм представляет собой своего рода сожительство. С ним тесно переплетаются другие формы сожительства животных, растений и животных с растениями, возникающие также на основе совместного обитания и сходного питания.
Иногда какие-нибудь сидячие формы животных поселяются на теле других организмов, но могут жить и на других предметах. Такие временные пространственные связи не являются сожительством. Здесь налицо случайное поселение одного животного на другом. При этом устойчивых отношений между видами не возникает.
Если сожительство не носит случайного характера и сочетание двух и более организмов систематически повторяется, то между видами образуются определенные формы биотических связей. Различают несколько типов таких сожительств.
Комменсализм (нахлебничество, сотрапезничество) – взаимоотношения на базе пищевых связей, при которых один из партнеров извлекает выгоду, а для другого они безразличны. При комменсализме обычно мелкие организмы, поселяясь возле или внутри крупных, пользуются не только территорией, но и остатками пищи последних. Так, краб пинникса (Pinnixa), поселяющийся в раковине дальневосточных пластинчатожаберных моллюсков (Pecten jessoensis), питается их объедками. Мальки многих рыб живут среди щупалец актиний и медуз и питаются отбросами их пищи. Многие птицы (жаворонки, овсянки, воробьи, даже куропатки) кормятся на экскрементах копытных, выбирая оттуда непереваренные зерна растений. Песцы в тундре следуют за белым медведем и доедают остатки его пищи. Часто питание комменсалов становится вредным для партнера. Тогда комменсализм граничит с паразитизмом.
Синойкия, или квартирантство,– пространственное сожительство, полезное для одного вида и безразличное для другого. Иногда между комменсализмом и синойкией трудно установить различия. При синойкии, как правило, не возникает непосредственных пищевых отношений. Например, мелкие организмы, поселяются на крупных, используя последних как субстрат. Это рыба-прилипала – на теле акулы, кишечнополостные – на раковинах моллюсков, усоногие раки – на панцирях крабов, на коже китов и акул. Такое поверхностное размещение мелких организмов на крупных называется эпойкией. Когда мелкие организмы размещаются внутри крупных, это будет энтойкия. Примером может служить рыба фиерасфер (Fierasfer), живущая в водных легких голотурий (Holothuria tubulosa).
Часто мелкие, слабоподвижные животные используют крупных для расселения. Некоторые мучные клещи (Tyroglyphoidea), например, временно прикрепляются к телу более подвижных насекомых или даже мышей и крыс и с их помощью переселяются в новые места обитания. Такая форма синойкии получила название форезия.
К синойкии в растительном мире можно отнести и эпифитность. Эпифиты поселяются на поверхности тела других растений. Это многие лишайники и мхи, растущие на стволах деревьев. Среди высших растений эпифитов особенно много в тропических лесах. Несмотря на тесную связь между эпифитами и растениями, на которых они поселяются, какого-либо активного влияния их друг на друга не обнаружено.
Взаимоотношения типа синойкии широко распространены у животных с растениями. Использование животными растительного субстрата для постройки жилищ (гнезда птиц, обитание в дуплах, в расщелинах коры, в свернутых листьях), перенос семян и плодов растений животными – все это примеры различных форм синойкии. Некоторые виды насекомых обитают в муравейниках и термитниках. Правда, ни муравейник, ни термитник не являются организмами, а служат жилищами, внутри которых поселяются и другие виды. Нередко синойкия становится начальным этапом более тесных взаимоотношений типа паразитизма и симбиоза.
Симбиоз, или мутуализм,– обоюдовыгодное сожительство разных видов. Классический пример симбиоза – сожительство рака-отшельника (Pagurus bernhardus) и актинии (Sagartia parasitica). Оба вида получают взаимную пользу от такой связи. Форму симбиоза приобретают отношения многих муравьев и тлей: муравьи защищают тлей от врагов, а сами питаются их сахаристыми выделениями.
Наблюдается симбиоз и среди растений. Широко распространенные лишайники – не что иное, как симбиоз между водорослями и грибами. Симбиотические отношения высших растений с грибами представляет собой микориза. При эндотрофной микоризе гифы гриба проникают в клетки корня растения и питаются как паразиты. А высшие растения, частично усваивая гифы, получают дополнительное азотистое питание и в свою очередь подкармливают гриб продуктами фотосинтеза (рис. 29). Взаимная польза от подобного сожительства настолько велика, что многие высшие растения (орхидные, вересковые, некоторые злаки) не могут расти без грибов, а грибы без этих растений.
При экзотрофной микоризе, характерной для большинства кустарниковых и древесных пород, гифы грибов лишь плотно прилегают к их корням, не проникая в клетки. Грибы пользуются выделениями корней и одновременно стимулируют усвояемость высшими растениями органических веществ почвы. Грибы и многие деревья и кустарники не могут жить друг бее друга.
Следует отметить, что комплекс отношений типа симбиоза содержит в себе самые разнообразные переходы – от отношений более или менее индифферентных до таких, когда оба члена сожительства обеспечивают взаимное существование.
Зоохория – одна из форм межвидовых взаимоотношений, заключающихся в распространении животными семян и спор растений. При этом как у животных, так и у растений вырабатывается ряд специфических приспособлений.
При эктозоохории семена, прикрепившись с помощью различных зацепок, крючков, щетинок, клейких веществ к шерсти млекопитающих, к перьям птиц, к телу насекомых, разносятся ими на далекие расстояния. Отрываясь в различных местах, при наступлении благоприятных условий они прорастают. Нередко мелкие семена пристают к телу животных с грязью, а когда она, высохнув, стряхивается, то вместе с ней отваливаются и семена. Так распространяются многие степные, луговые, прибрежные и водные растения.
Еще более тесные взаимоотношения между животными и растениями складываютя при эндозоохории. Животные поедают ягоды, сочные плоды, и семена, содержащиеся в них, не только не перевариваются, а даже, как установлено, под воздействием желудочного сока повышают свою всхожесть. Вместе с экскрементами они выделяются наружу, оказываясь, таким образом, в самых различных местах.
Кроме того, было обнаружено, что в процессе жизнедеятельности растения, животные и микроорганизмы выделяют во внешнюю среду газообразные, жидкие и твердые вещества (фитонциды), которые, обладая определенной биологической активностью, оказывают влияние на другие организмы. Явление это известно с давних пор, но лишь в 1937 г. немецкий ботаник X. Молиш дал ему название аллелопатии. Аллелопатия – это взаимодействие организмов посредством специфически действующих химических продуктов обмена веществ. Наиболее глубоко данное явление изучено на растительных организмах. Выделения растений оказывают на организмы сообщества либо токсическое, либо стимулирующее действие. Так, выделения соплодий свеклы тормозят прорастание семян куколя (Agrostemma githago). Абсинтин листьев полыни горькой (Artemisia absinthium) угнетает развитие многих растений. Нут (Cicer arietinum) подавляюще действует на картофель, томаты, кукурузу, подсолнечник и другие культуры; фасоль – на рост яровой пшеницы; корневые выделения пырея (Agropyron repens) и костра (Bromus inermis)–на растущие вблизи с ними другие травянистые растения и даже деревья.
На животных растения оказывают либо аттрактивное (привлекающее), либо репеллентное (отталкивающее) действие. Это особенно важно для вредителей при выборе кормовых растений и для паразитов при отыскивании растений-хозяев. К примеру, ослабленные в результате физиологических нарушений деревья выделяют летучие вещества, информирующие стволовых вредителей о том, что они могут их заселять. Аттрактивное действие таких соединений наиболее сильно проявляется у свежесрубленных деревьев, т. е. у умирающих растений, находящихся на пределе физиологической ослабленности. Немало известно и отпугивающих, а также ядовитых для фитофагов веществ, лимитирующих их распространение в сообществе.
Кроме названных, имеются многочисленные колины и другие растительные выделения, специфически действующие на другие виды растений.
Животные выделяют феромоны (телергоны) – своеобразные активные вещества, влияющие на развитие и поведение индивидуумов своего вида, а также сообщающие определенную информацию другим видам.
Биологически активные вещества вырабатывают и многие микроорганизмы. Широкую известность получили, например, антибиотики пенициллин, стрептомицин и другие продукты жизнедеятельности грибков рода Penicillium и Streptomyces. Большой популярностью пользуется также гибберилин грибка Gibberella, стимулирующий рост высших растений.
Взаимоотношения, при которых возникают отрицательные условия для одной или нескольких популяций, получили название аменсализм. Аменсализм можно рассматривать как одну из форм антибиоза или как крайнюю форму аллелопатии (невозможность существования того или иного вида в присутствии другого в результате интоксикации среды). Таковы, например, взаимоотношения плесневых грибов с бактериями.
Наиболее сложные биотические связи возникают у млекопитающих, птиц и особенно у насекомых. Так, муравьи-амазонки (Polyergus rufescens) совершают набеги на чужие муравейники, захватывают там личинок и куколок и выводят из них в своем муравейнике взрослых муравьев. Они и выполняют всю работу по уходу за яйцами, личинками, куколками и плодущей самкой, по уборке и достройке жилища муравьев-амазонок. Муравьи россомирмексы (Rossomyrmex proformicarurn) также выводят в своем муравейнике куколок степного медового муравья (Proformica epinotalis). Харпагоксе-нус (Harpagoxenus) похищает куколок дернового муравья (Tetramorium caespitum). Эта своеобразная форма межвидовых взаимоотношений названа рабовладельчеством. Муравьи-рабовладельцы обитают только в странах северного полушария с умеренным климатом. По всей вероятности, такая форма сложных межвидовых взаимоотношений возникла на базе материнского инстинкта, поскольку установлено, что многие муравьи, не будучи рабовладельцами, при набеге на муравейники других видов не уничтожают куколок и личинок, а приносят их в свой муравейник и выращивают из них взрослых особей. «Рабовладельческий» инстинкт поэтому можно рассматривать как доведенный до крайности материнский инстинкт при хищническом нападении на чужую общину.
В естественных условиях в биоценозах, кроме рассмотреных межвидовых взаимоотношений, возникают различные переходные формы – от чисто антагонистических до подлинной взаимопомощи.
Влияние растений друг на друга. Каждый вид растения приспособлен к существованию в определенном биогеоценозе в условиях взаимовлияния всех членов сообщества. Взаимное воздействие растений друга на друга чрезвычайно разносторонне и зависит от многих факторов: от степени их скученности, от изменений во внешней среде (свет, влага, состав почвы, химизм, накопление отмершего органического вещества), вызываемых ими в процессе жизнедеятельности, от наличия питательных веществ.
При рассмотрении типов биотических отношений в биоценозе уже давались характеристики их форм. Кроме того, они подразделяются на следующие виды взаимоотношений.
Контактные взаимодействия (контактные коакции) возникают при соприкосновении, срастании и проникновении одних растений в другие. Они бывают механическими и физиологическими.
При механическом соприкосновении (касании) возможны искривление стволов, изменение формы кроны. Давление друг на друга оказывают корни растений, дерновины злаков: лианы воздействуют на опорные деревья. В лесу при ветре происходит схлестывание ветвей. В результате такого механического влияния растения в биоценозе размещаются на каком-то определенном расстоянии друг от друга.
Физиологические контакты возникают при паразитизме, когда растение-паразит и растение-хозяин взаимно влияют на те или иные функции друг друга, а также при симбиозе (микоризы, лишайники, бактериоризы). У хозяина при этом вырабатываются такие защитные приспособления физиологического порядка, как реакция клеточного сока, выделение ферментов, препятствующих проникновению паразита в клетки и ткани, и многие другие. У паразитов также формируются физиологические особенности. Они направлены на подавление защитных реакций хозяина. При заражении хозяин становится питательной средой паразита, и по мере размножения последнего все сильнее испытывает его влияние.
При микоризе возникают столь тесные связи партнеров, что они друг без друга существовать не могут. Орхидные (Orchidaceae), вересковые (Ericaceae), грушанковые (Pyrolaceae) и другие микотрофы способны нормально развиваться только при наличии микоризных грибов. А такие высшие растения, как подъельник волосистый (Monotropa hypopitys) и петров крест (Lathraea squamaria) полностью перешли на микротрофное питание и утратили хлорофиллоносные органы. Это яркий пример контактного физиологического взаимодействия растений, доведенного до крайности.
Лишайники (Lichenophyta) –это своеобразные симбиотические организмы. Тело их (слоевище) состоит из двух компонентов – грибов и водорослей,– находящихся в состоянии взаимно выгодного физиологического мутуализма или, точнее, обоюдного паразитизма (алело-паразитизма). Грибы доставляют водорослям влагу и растворенные минеральные вещества, а водоросли перерабатывают их в органические соединения, обеспечивая питание грибов и свое собственное. Лишайники чрезвычайно устойчивы к неблагоприятным условиям, живут и размножаются там, где в отдельности их симбионты погибают: в тундре, в альпийских и субальпийских каменистых пустынях. В то же время они очень чувствительны к чистоте воздуха и не выносят даже незначительного его загрязнения дымом, копотью и особенно сернистым газом.
При бактериоризах возникает прочная связь корней растений с бактериями. Бактерии, питаясь органическими веществами корневых выделений растения-хозяина, отдают ему свои продукты жизнедеятельности (минеральные и органические соединения, витамины). Часто бактерии проникают в клетки корня и на нем образуются клубеньки. Довольно глубокие отношения устанавливаются между клубеньковыми бактериями и бобовыми растениями. Они друг без друга хуже растут и плохо развиваются.
Косвенные взаимодействия растений осуществляются не при непосредственном их контакте, а через изменение абиотической и биотической среды. Они значительно сложней и разнообразней контактных взаимодействий. При таких трансабиотических коакциях взаимовлияние растений происходит через ухудшение или улучшение абиотической среды обитания. Например, ослабление либо гибель какого-то растения в результате поражения его паразитами или вследствие вырубки, пожара, наводнения позволяет соседним занять освободившееся место; грибная или бактериальная флора на корнях одного растения изменяет химические и физические свойства почвы, делая ее пригодной для одних растений и недоступной для других; многие виды растений, как известно, выделяя колины, фитонциды, спирты, эфиры, кислоты, алкалоиды, создают аналогичный эффект.
Во взаимоотношениях растений можно обнаружить все переходы от взаимно полезных до вредных. Вырабатывающиеся в процессе эволюции адаптации растений друг к другу обусловливают распределение их в биоценозах, а также влияют на структуру и функциональную деятельность фитоценозов. При этом необходимо учитывать, что характер взаимосвязей растений меняется во времени и это приводит к перестройке биоценозов, к сезонной и годичной смене растительности. Например, сближенность взрослых деревьев в лесу полезна каждому виду лишь до известного предела и при строго определенных условиях. Семена многих растений при посеве кучками (гнездовой посев) значительно лучше прорастают, чем одиночные. Но позже сближенное расположение поростков отрицательно влияет на рост: растениям становится тесно.
Особенно сильно проявляется взаимное воздействие растений в самых разнообразных формах в смешанных многовидовых сообществах. Существеннейшим образом отличаются деревья, выросшие в густом лесу и на опушке. Это касается и травянистой растительности, а также кустарников. В природе наиболее жизнеспособны многовидовые фитоценозы. Условия, складывающиеся в них, благоприятствуют процветанию каждого вида, входящего в его состав, хотя может случиться, что растения, выросшие одиночно, выглядят лучше, чем выросшие в лесу или степном разнотравье (см. рис. 7).
Довольно сложные взаимоотношения между культурными растениями и сорняками складываются в агробиоценозах. Здесь преобладает их взаимное угнетение. Вместе с тем хорошо известно, что смешанные посевы (различные травосмеси в луговодстве, смешанные посадки леса, викоовсяные и другие злаково-бобовые смеси) более продуктивны, чем чистые (одновидовые), так как могут полнее использовать среду. Именно этим объясняются проблемы, возникающие при создании лесных монокультур, т. е. чистых однопородных насаждений. Естественные биоценозы обычно представляют собой смешанные, т. е. многовидовые сообщества. И чем разнообразнее по видовому составу биоценоз, тем у него больше возможностей для полного и экономичного освоения условий жизни. Одновременно в многовидовом фитоценозе формируются чрезвычайно сложные взаимовлияния растений, направленные в конечном итоге на выработку самых выгодных для биогеоценоза количественных и качественных соотношений между видами.
Жизнь на Земле могла развиваться только как особая форма круговорота веществ и потока энергии. Круговорот веществ осуществляется в первую очередь на основании деятельности большого разнообразия организмов.
Каждый организм потребляет из окружающей среды необходимые для жизнедеятельности вещества, а возвращает уже непригодные. Причем некоторые виды потребляют нужные им вещества непосредственно из среды, другие используют продукты, переработанные и выделенные первыми, третьи – вторыми и так до тех пор, пока вещество снова не превращается в свое первоначальное состояние и не попадает во внешнюю среду. Отсюда и возникает необходимость сосуществования различных организмов (видовое многообразие), способных использовать продукты жизнедеятельности друг друга. На основе такого сосуществования и формируются разнообразные биотические связи.
Биотические связи животных и растений. Растения и животные, автотрофы и гетеротрофы, с первых дней возникновения развиваются совместно, обусловливая возможность своего сосуществования. С древнейших времен и до настоящего момента они проходят свой эволюционный путь в тесном контакте, в тесной взаимосвязи и взаимозависимости. В процессе сопряженной эволюции у различных видов растений и животных выработались взаимные приспособления друг к другу, коадаптации, столь прочные, что раздельно жить в современных условиях они уже не могут. В этом проявляется единство органического мира. О связи растительных и животных организмов в биоценозе писал еще в 1912 г. Г. Ф. Морозов: «...жизнь животных в лесу тоже тесно спаяна со свойствами леса... тоже находится в зависимости от леса, с одной стороны, а с другой стороны, влияет на жизнь леса. В лесу стихийном, или девственном, нет ни полезных, ни вредных животных – там все полезны для леса; и белка, и сойка, и другие животные, поедающие желуди, не являются, однако, в силу этого истребителями дуба, а, наоборот, агентами распространения его, факторами жизни и распространения леса. Лес есть не только общежитие древесных растений, он представляет собою общежитие более широкого порядка; в нем не только растения приспособлены друг к другу, но и животные к растениям, и растения к животным; все это взаимно приспособлено друг к другу».
Примером исторически возникших глубоких взаимных приспособлений могут служить коадаптации насекомоопыляемых растений и насекомых-опылителей, орнитофильных растений и их опылителей птиц, зоохория. Животные, и прежде всего насекомые, птицы и млекопитающие оказали огромное влияние на растения, на развитие у них ярко окрашенных цветков, на формирование защитных приспособлений анатомо-морфологического, биохимического и физиологического характера, на эволюцию съедобных плодов.
С другой стороны, растения также сыграли большую роль при взаимоотношениях с животными. Именно в процессе сопряженной эволюции определились группы животных со специализированным питанием: плодоядные, семеноядные, питающиеся нектаром, зелеными органами растений, и т. д. Результатом совместной эволюции животных и растений является и приуроченность различных групп животных к определенным группам растений.
Цветковые растения появились в меловой период. Как отмечает С. С. Станков, обилие солнечных лучей, яркие и своеобразные цветки и армия насекомых вывели цветковые растения на первое место. Вместе с ними развились и насекомые-опылители – процветающая в настоящее время группа животных. Многие современные животные особенно тесно связаны с покрытосеменными растениями. Именно вместе с ними шла их эволюция. По мнению М. И. Голенкина, возникновение покрытосеменных было настоящей и глубокой революцией на Земле, той гранью, начиная с которой жизнь не только растений, но и вообще всех живых существ пошла совершенно иным путем.
Тесная взаимозависимость растений и животных выражается и в том, что растения в процессе исторического развития приспособились существовать в условиях обязательных потерь, связанных с прокормом фитофагов путем избыточного продуцирования, рассчитанного на эти потери. Фитофаги же, поедая часть зеленой массы растений, ускоряют круговорот веществ и перевод органического вещества в минеральное. Минеральные вещества попадают в почвенные растворы и снова используются растениями.
К сожалению, данных, позволяющих четко проследить исторические связи животных и растений, сравнительно немного. Тем не менее существование в природе различных биогеоценозов, к примеру группировок растений и животных, типичных для степей и пустынь, для хвойных и смешанных лесов, служит неоспоримым свидетельством этих связей. В каждом биогеоценозе между растениями и животными устанавливаются разнообразные биотические связи, обеспечивающие развитие этого биогеоценоза во времени и пространстве.
Трофические связи животных и растений. Во всем многообразии биотических взаимоотношений растений и животных, как уже неоднократно отмечалось, первостепенное значение имеют трофические. Это так называемые взаимоотношения автотрофных и гетеротрофных организмов. Растения служат пищей для большой группы растительноядных животных – фитофагов и тем самым определяют их численность, географическое распространение и распределение по местам обитания. К фитофагам можно отнести и большую группу организмов, существующих за счет незеленых, бесхлорофилльных растений. Практически все организмы, за исключением хищников и паразитов, питаются растительной пищей. Правда, есть еще специализированная группа сапрофагов, живущих за счет разлагающегося органического вещества, а также копрофагов, питающихся экскрементами животных. Но между сапрофагами и фитофагами иногда трудно обнаружить различия, ибо сапрофагия часто начинается там, где еще не окончена типичная фитофагия.
Пищевая специализация растительноядных животных чрезвычайно разнообразна. Имеются типичные сосущие фитофаги, которые высасывают соки растений. Многие питаются вегетативными, зелеными, частями их, обгрызая листья, побеги, почки. Это млекопитающие, птицы, моллюски, насекомые. Ряд животных кормится подземными частями растений, повреждая корни, луковицы, клубни. При всем этом многообразии одни животные уничтожают растение целиком, другие объедают его частично, третьи внедряются внутрь стволов, стеблей, листьев и ведут там паразитический образ жизни.
В воде мелкие животные (зоопланктон), а также более крупные формы на различных стадиях развития поедают зеленые водоросли (фитопланктон). Рыбы, моллюски, членистоногие, водные млекопитающие, водоплавающие птицы употребляют в пищу водные растения. Попугаи-лори (Trichoglossidae), нектарницы (Melipha-gidae), колибри (Trochilidae) из птиц, некоторые летучие мыши (Phyllostomidae) из млекопитающих и особенно насекомые питаются нектаром и пыльцой цветков. Фитофагами являются также животные, питающиеся ягодами, сочными плодами и семенами растений.
В определенные периоды жизни растительную пищу употребляют и типичные плотоядные животные. Так, соболь и куница едят ягоды и семена растений; синица, дятел и ряд других насекомоядных птиц – семена деревьев и трав. Землеройки в зимний период часто переходят на растительные корма. Трудно среди животных найти виды, которые бы совершенно не питались растительной пищей.
На базе таких пищевых взаимоотношений возникли специализированные группы животных, приспособившиеся жить за счет определенных растений. Так, травянистой растительностью питаются травоядные животные. Это в основном обитатели степных ландшафтов: копытные, многие грызуны, перепела, куропатки, насекомые. Все степные формы адаптировались к жизни на открытых пространствах, к питанию в основном грубыми кормами. Они имеют острое зрение, быстро бегают, им свойственно особое строение пищеварительной системы.
В связи с приуроченностью к тому или иному виду пищи у различных групп животных выработались морфо-физиологические и экологические признаки, характеризующие каждый вид и его пищевую специализацию. Питание трудноперевариваемой пищей, в частности клетчаткой, привело к удлинению кишечника, а у многих животных появились дополнительные отделы его. Так сформировался сложный желудок жвачных, где вдобавок ко всему поселяются инфузории-симбионты (Entodi-niomorpha), способствующие усвоению клетчатки. Подобную роль играют симбионты-жгутиковые в кишечнике термитов, питающихся мертвой древесиной.
Специализированную
группу фитофагов образуют лесные
животные. Например, клесты (Loxia),
употребляющие семена хвойных пород,
имеют особое строение клюва и поэтому
распространены только там, где есть
достаточное количество кормовых
растений.

При этом в зависимости от преобладания в пище сосновых, еловых или пихтовых семян строение клюва изменяется (рис. 30). С ареалом кедровой сосны (Firms sibirica) связано распространение кедровки (Nucifraga caryocatacles). И клесты, и кедровки, так же как и многие другие животные, в случае неурожая кормовых растений или при уничтожении последних конкурирующими видами вынуждены совершать далекие миграции в поисках пищи. Во время таких миграций часто наблюдается их массовая гибель. Недостаток пищи обычно вызывает колебания численности животных-фитофагов. В этом особенно проявляется зависимость животных от растений.
Поскольку многообразие зеленых растений дает животным-фитофагам разнокачественный корм, у последних исторически выработалась пищевая специализация, появились стенофаги и эврифаги, а также всевозможные переходные между ними формы (монофаги, олигофаги, полифаги). Монофагов в природе немного, и все они прочно связаны со своими кормовыми растениями. При неурожае или гибели кормового растения монофаги могут полностью погибнуть, несмотря на то, что узкоспециализированные в пищевом отношении животные обладают способностью совершать миграции или переносить длительные голодовки.
В более выгодном положении оказываются животные, приспособившиеся жить за счет нескольких (олигофаги) или многих видов (полифаги) растений. Такие животные в случае исчезновения основного корма могут переключаться на новые растения.
Для ряда животных причиной миграций и других форм сложного поведения является сезонная смена кормовых запасов. При этом многие из них с наступлением холодов перемещаются на юг не потому, что не могут переносить низкие температуры, а в связи с исчезновением их обычного корма, в поисках которого они порой вынуждены преодолевать огромные расстояния. Конечно, не все миграции обусловлены изменениями кормовой базы. Но в основном массовые перемещения животных вызваны именно этим фактором.
Сезонные изменения запасов и качества растительного корма влияют и на поведение, а также на образ жизни животных-фитофагов. Суслики, хомяки, хомячки и многие другие на зиму впадают в спячку, первопричиной которой служит исчезновение основных растительных кормов.
У насекомых-фитофагов умеренного климата к осени при понижении температуры, сокращении светового дня и прекращении вегетации растений (исчезновение корма) развитие приостанавливается на весь зимний период. Наступает прекращение питания (еще до появления отрицательных температур осенне-зимнего периода), диапауза, которая служит своеобразной пищевой адаптацией.
Зависимость географического распространения фитофагов от ареала их кормовых растений особенно наглядно проявляется у насекомых. Тополевый (Melasoma populi) и осиновый (М. tremulae) листоеды, например, встречаются лишь там, где растет осина; ольховый листоед (Agelastica alni) связан с ареалом ольхи серой (Alnus incana); рыжий и обыкновенный сосновые пилильщики (Neodiprion sertifer, Diprion pini) обитают только в сосновых насаждениях, а капустные белянка и совка – в районах произрастания крестоцветных растений (Cruciferae), в том числе и качанной капусты. Однако приуроченность фитофагов к кормовым растениям не абсолютна. Так, данные И. К. Лопатина свидетельствуют, что тополевый, осиновый и ольховый листоеды в Средней Азии и Казахстане живут на ивах, поэтому ареалы, в частности ольхового листоеда, гораздо шире, чем ольхи.
Процессом распространения и распределения растительного и животного мира на Земле можно управлять. Прокладывание, к примеру, лесозащитных полос в степной полосе обеспечивает продвижение с севера на юг типичных лесных обитателей – птиц, зверей, насекомых. С другой стороны, уничтожение лесов – вырубка, раскорчевка – при последующей распашке этих площадей сокращают ареалы лесных животных и способствуют проникновению в лесные зоны степных обитателей зайца-русака (Lepus europaeus), хомяка (Cricetus cricetus), перепела (Coturnix coturnix), полевого жаворонка (Alauda arvensis), серой куропатки (Perdix perdix), ряда насекомых. Таким образом, расширение посевных площадей в степной и лесной зонах, создание лесных полос в степи изменяют растительный покров, а это влечет за собой изменение в животном мире, в первую очередь среди животных-фитофагов.
Кроме фитофагии, в природе распространены и диаметрально противоположные взаимоотношения – зоофагия, т. е. питание растений животными. Группу типичных зоофагов составляют насекомоядные растения. Их в современной флоре насчитывается до 500 видов. Это росолистник (Drosophyllum lusitanicum), венерина мухоловка (Dionaea muscipula), альдрованда (Aldrovanda vesiculosa), непентес (Nepenthes), дарлингтония (Dar-lingtonia), пузырчатки (Utricularia) и многие другие. Все эти растения имеют самые различные приспособления для ловли насекомых. Так, у саррацении (Sarracenia purpurea), распространенной в Америке, листья свернуты в трубку, на дне которой собирается жидкость, привлекающая насекомых. Попадая в эти трубки, насекомые уже не могут выбраться оттуда и становятся добычей растения – оно их переваривает. Своеобразное приспособление образовалось для ловли мелких насекомых и ракообразных у водного растения пузырчатки. При соприкосновении этих животных с чувствительным волоском пузырька последний открывается и жертва вместе с водой с силой всасывается в него. После этого пузырек сразу же закрывается и растение начинает переваривать добычу. В умеренных широтах на торфяных сфагновых болотах, бедных азотом и минеральными солями, часто встречаются насекомоядные растения росянки (Drosera). Питаясь насекомыми, росянки не испытывают ни азотного, ни фосфорно-калийного голодания. Некоторые грибы ловят почвенных нематод с помощью микроскопических силков или клейких утолщений, но это уже растения-зоофаги из мира бесхлорофилльных, нефотосинтезирующих растений.
На основе трофических связей животных с растениями в природе широко распространены паразитические взаимоотношения. Среди типичных паразитов растений можно назвать многих насекомых, клещей, высасывающих соки, галлообразователей. Некоторые из них служат переносчиками возбудителей болезней растений. Личинки ряда мух, жуков, бабочек, внедряясь в стволы, стебли, листья и другие органы растений, паразитируют как эндопаразиты. Подземные части растений поражаются растительноядными нематодами, личинками насекомых.
Однако паразитируют не только животные на растениях, но и растения на животных. К последним прежде всего относится большая группа паразитических грибов, бактерий, являющихся возбудителями тех или иных болезней.
Роль животных в опылении растений. В процессе эволюции у растений изменялся способ оплодотворения. В начале развития наземных растений для слияния половых клеток требовалась вода. Затем у голосеменных водная среда заменилась воздушной. На следующем этапе, с появлением покрытосеменных, опылителями в большинстве случаев становятся животные, в основном насекомые. В настоящее время в Европе до 80 % видов покрытосеменных растений опыляется насекомыми, 19 % – ветром и около 1 % другими способами. Собирая нектар в различных цветках, насекомые разносят пыльцу и тем самым способствуют оплодотворению растений. При этом они обеспечивают воспроизведение только тех растений, у которых в момент цветения имеются излишки нектара. Эти растения в процессе естественного отбора широко расселились, вместе с ними распространились и насекомые-опылители. В результате и те и другие оказались в тесной взаимозависимости.
Среди опылителей, кроме насекомых, много птиц. Растения, опыляемые птицами, называются орнитофильными. Всего насчитывается до 1600 видов птиц-опылителей. Это колибри (Trochilidae) и цветочницы (Coerelidae) в Америке, медососы (Meliphagidae) и попугаи-лори (Loriinae) в Австралии, нектарницы (Necta-riidae) и цветососы (Dicaeidae) в Африке и др.
Есть опылители и среди других животных. Многие древесные породы в лесах Южной и Центральной Америки, например, опыляются исключительно летучими мышами из семейства листоносых (Phyllostomidae). Опыление растений летучими мышами получило название хироптерофилии. Считается, что распространенное в умеренных широтах растение селезеночник (Chrysosplenium alternifolium) в основном опыляется улитками (малакофилия), хотя достоверных доказательств этого пока нет.
У специфичных опылителей – эвтропных насекомых – развились очень тонкие приспособления, направленные на обеспечение перекрестного опыления цветковых растений. Эти приспособления столь глубоки и причудливы, что по сей день остаются загадками природы. Вместе с тем специфика данных взаимоотношений строится прежде всего на обоюдной выгоде.
Однако тесная взаимосвязь и взаимная приспособленность характерна не для всех видов опылителей и растений. Строение цветка той или иной группы растений позволяет проникнуть к нектару лишь определенным насекомым. Этим обусловливается равномерное распределение опылителей и перекрестное опыление растений. Примером такой специализации является развитие на одном растении мужских и женских цветков, которые достигают половой зрелости неодновременно. У ряда растений тычинки и пестики созревают в различное время (дихогамия). Интересны растения, имеющие цветки двух форм: для одних характерны длинные тычинки и короткие пестики, у других, наоборот, тычинки короткие, а пестики очень длинные (рис. 31). Такое явление получило название гетеростилии. Значение его в том, что насекомые у таких цветков могут производить лишь перекрестное опыление; самоопыление же в пределах одного цветка полностью исключается.

Таким образом, разнообразное и сложное строение цветков – различная форма лепестков, симметричное или несимметричное их расположение, наличие определенных соцветий – все это приспособления к строению тела и поведению строго конкретных насекомых. Например, цветки дикой моркови (Daucus саrоta), тмина (Carum carvi) опыляются муравьями. Более же крупные цветки смолки (Viscaria vulragis) муравьи опылить не могут.
Исключительно тесные и прочные связи выработались у юкки (Jucca gloriosa) и опыляющей ее юкковой моли (Pronuba juccasella). Самка моли щупиками собирает пыльцу, скатывает ее в шарик и, держа его над головой, подлетает к другому цветку. Здесь она при помощи яйцеклада откладывает яйцо в столбик пестика, а затем в углубление на рыльце аккуратно укладывает принесенный шарик пыльцы. Развивающаяся из яйца личинка питается семенами юкки. Если бабочка не положит на рыльце пестика пыльцу, семена не будут развиваться и личинка погибнет. С другой стороны, юкка продуцирует такое большое количество семян, что личинка, уничтожая часть их, не наносит ущерба растению. Такое интересное и сложное инстинктивное поведение юкковой моли обеспечивает и питание ее личинки и плодоношение юкки.
При этом различные виды моли способны опылять строго определенные виды юкки. Подобные взаимоотношения сложились между некоторыми сортами культурного инжира (Ficus) и дикими видами его с хальцидом бластофагой (Blastophaga psenes).
Исключительное значение как опылители имеют общественные пчелы. Приведем такой пример. В среднем семья шмелей состоит из 100 рабочих особей, совершающих в день не менее чем по 20 вылетов. За каждый вылет они посещают приблизительно по 240 цветков, а за день – по 4800. Если учесть, что рабочий шмель живет до одного месяца, то выходит: за месяц вся семья может опылить 10–12 миллионов цветков. Но семья шмелей существует значительно больше месяца, поскольку по мере гибели одних рабочих особей их место занимают молодые. Еще более внушительные цифры обнаруживаются при оценке опылительной деятельности пчелиной семьи. Установлено, что рабочая пчела за минуту облетает 12 цветков, а за день – около 7200. В слабой семье бывает до 10 000 рабочих особей, а в сильной – до 50 000. Даже по приблизительным подсчетам пчелы одной семьи в день опыляют не менее 360 миллионов цветков.
Таким образом, связи насекомых-опылителей с цветковыми растениями, развившиеся в течение длительной эволюции, постепенно выразились в такой тесной взаимной зависимости, что раздельно их существование невозможно.
Однако не все насекомые, посещающие цветки, способны их опыливать. Большинство муравьев, некоторые жуки лишены этого свойства. Такие насекомые называются дистрοпными. Дистропия обусловлена тем, что у ряда растений в процессе эволюции выработались приспособления, препятствующие проникновению некоторых насекомых в их цветки. Иногда дистропия связана с поведением самих насекомых, которые все же изредка переносят пыльцу с цветка на цветок. Это так называемые аллотропные насекомые. К ним относятся, например, мухи, питающиеся нектаром.
Роль животных в распространении растений. На основе трофических и пространственных связей растений и животных сформировались определенные взаимоотношения их, названные зоохорией. (Частично зоохория была рассмотрена как форма межвидовых взаимоотношений.) Зоохория – это распространение животными семян и спор растений. Различают экто- и эндозоохорию.
Эктозоохория бывает активной и пассивной.
Главную роль при активной эктозоохории выполняют животные. Они, делая запасы пищи, собирают семена и переносят их на значительные расстояния. Семена, которые переносят звери, птицы, насекомые, обычно крупных размеров и без специальных приспособлений для прикрепления к телу животного. Поэтому при транспортировке часть семян, как правило, теряется.
Разные животные в зависимости от специфики питания распространяют определенные растения. Так, кедровка (Nucifraga caryocatactes) разносит в основном семена кедровой сосны (Pinus sibirica). Большие запасы кедровых орешков делает и бурундук (Eutamias sibiricus). Однако не все семена используются в пищу либо из-за гибели зверьков, либо из-за потери. Это и способствует прорастанию уцелевших семян в местах, подчас весьма далеких от растения, которому они принадлежали.
В распространении лесного орешника лещины (Соrуlus avellana) очень деятельны белки (Sciurus vulgaris). Они растаскивают орехи по всему лесу. В результате лещина расселяется на огромных площадях. Мелкие мышевидные грызуны также запасают много семян на зиму. Подсчитано, что одна пара желтогорлых мышей (Apodemus flavicollis) уносит в день до 5000 буковых орешков, а за две недели до 38 000. Немалую роль в расселении дуба (Quercus robur) играют сойки (Carrulus glandarius). Описан случай, когда в воронежских лесах на одном гектаре сосняка, расположенного в нескольких километрах от дубравы, было обнаружено свыше 500 молодых дубков, выросших из желудей, принесенных сойками. Запасают желуди на зиму и грызуны. Причем масштабы их деятельности в этом довольно велики. Лесная мышь (Apodemus sylvaticus), например, за 6 дней может утащить до 1500 желудей, а самец желтогорлой мыши (A. flavicollis) такое же количество запасает за 15 дней. Часть из собранных желудей зверьками не используется.
Распространению ряда растений способствуют также муравьи. Они накапливают около муравейников семена чистотела (Chelidonium), фиалок (Viola), медуницы (Pulmonaria), хохлатки (Corydalis), марьянника (Melampyrum), чистяка (Ficaria), споры грибов. Подсчитано, что одна колония муравьев Formica rufa рассеивает на расстоянии до 70 м от муравейника свыше 36 000 семян различных растений. Многие из них имеют привлекающие муравьев приспособления в виде придатков ариллоидов (элайосомы), содержащих большое количество маслянистых веществ. Муравьи съедают только эти придатки, а сами семена на следующий год прорастают на некотором удалении от растения. Семена с ариллоидами имеются у копытня (Asarum europaeum), ивана-да-марьи (Melampyrum nemorosum), перелески (Hepatica nobilis), гусиного лука (Gagea) и других растений. Все они, как правило, плодоносят в середине лета, когда муравьи наиболее активны.
Пассивная эктозоохория заключается в том, что семена растений транспортируются на значительные расстояния, прикрепляясь с помощью специальных приспособлений к телу животных. Для этого, например, у гравилата (Geum) на каждом плодике есть крючок, на плодах ложносытевидной осоки (Carex pseudocyperus) образуются цепкие щетинки и крючки, у линией (Linnaea borealis) семена обладают клейкими железистыми волосками, а плоды недотроги (Impatiens nolitangere) собраны в коробочки, створки которых при прикосновении резко раскрываются, и семена выбрасываются на расстояние до 1,5 м. Аналогично со струей клейкой жидкости разлетаются на расстояние до 6 м и семена бешеного огурца (Ecballium elaterium). Нередко, как уже упоминалось, семена растений пристают к телу животных с илом, грязью и таким образом разносятся иногда на тысячи километров.
Еще более сложные коадаптации между растениями и животными формируются при эндозоохории. У большинства птиц и у многих млекопитающих семена, пройдя через пищеварительный тракт, сохраняют жизнеспособность и в результате транспортируются на довольно далекие расстояния. В Норвегии подсчитали, что со свежим конским навозом, вносимым на поля как удобрение из расчета 60 т/га, засевается на каждом гектаре до 900000 сорных растений. Так распространяются семена ярутки (Thlaspi arvense). Овцы, лошади и другие домашние животные разносят пастушью сумку (Capsella bursa-pastoris), овсюг (Avena fatua), горчицу (Sinapis), дикую редьку (Raphanus raphanistrum), мокрицу (Stellaria media). Воробьи разносят семена земляники, бузины, калины, крушины, жимолости и других ягодников. Большую роль в распространении растений играют перелетные птицы. Благодаря им растения северных областей попадают в степи. Это шиповник, черемуха, крушина и др. Дрозд деряба (Turdus viscivorus) переносит семена омелы. На Гавайских островах широко расселился сорняк лантана (Lantana camara) в связи с акклиматизацией там птицы майны (Acridotheres tristis). С деятельностью плодоядных птиц связывают появление большинства деревьев на океанических островах.
Иногда семена распространяются через несколько звеньев. К примеру, через рыбоядных птиц расселяются водные растения.
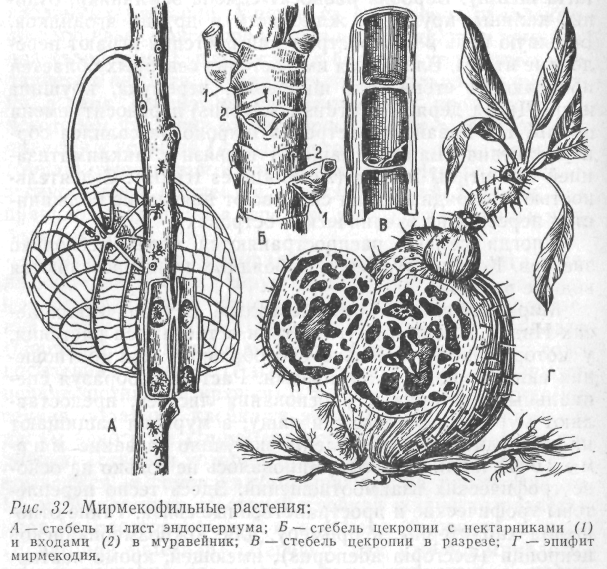
Мирмекофилия. «Грибные сады». В тропических лесах Индии, Китая и других стран встречаются растения, у которых исключительно своеобразные взаимоотношения складываются с муравьями. Растения, образуя специальные нектарники у основания листьев, предоставляют муравьям убежище и пищу, а муравьи защищают их от вредителей. Это явление получило название мирмекофилии. Оно сформировалось не только на основе трофических взаимоотношений. Здесь тесно переплетены трофические и пространственные связи. Такие отношения характерны, к примеру, для южноамериканской цекропии (Cecropia adenopus), имеющей, кроме нектарников, своеобразные полости внутри стебля, которые и заселяются муравьями. У эпифита мирмекодии (Myrmecodia pentasperma) вздутый стебель (ложная луковица) также пронизан полостями и представляет собой искусственный муравейник. У эпифита дисхидии (Dischidia rafflesiana) образуются листовые кувшинчики, где собирается влага, используемая самим растением. Некоторые из этих кувшинчиков оказываются перевернутыми вверх дном, и тогда их заселяют муравьи. У мексиканского кустарника Acacia cornigera муравьи поселяются в расположенных у основания листьев крупных полых колючках. Кроме того, на конце каждого листочка размещается особое тельце белого цвета, наполненное жиром и белком – прекрасным кормом для муравьев. Мирмекофильных растений в тропиках известно до 3000 (рис. 32).
К мирмекофилам относятся и многие животные, находящие в муравейниках укрытие и питающиеся отбросами муравьев, гниющим строительным материалом гнезда или превращающиеся в настоящих с и м φ и л о в, т. е. нахлебников или даже своеобразных паразитов и хищников муравьев. Известно свыше 2000 видов мирмекофильных членистоногих.
Между животными-мирмекофилами и муравьями формируются исключительно сложные взаимоотношения.
Симфилов муравьи кормят жидкой пищей, как и своих личинок. Симфилы же, благодаря наличию специфичных желез, выделяют вещества, которыми питаются муравьи. Мирмекофильными насекомыми являются, например, тропические пауссиды (Paussidae), жуки-ощупники (Pselaphidae), стафилиниды (Staphylinidae). Интересны взаимоотношения с муравьями у некоторых видов клещей семейства Antennophoridae. Они держатся на нижней стороне головы муравьев и слизывают с их щупиков и челюстей капельки пищи. Классическим примером мирмекофила является жук ломехуза (Lomechusa strumosa), встречающийся только в муравейниках. Иногда муравьи уделяют им так много внимания, что от этого страдают сами – семья их деградирует и нередко погибает. Сущность подобных взаимоотношений до конца еще не выяснена.
Группа беспозвоночных животных, ведущих аналогичный образ жизни в термитниках, получила название термитофилов.
Хотя биология мирмекофилов и термитофилов изучена слабо, все же форму связи их с муравьями можно охарактеризовать как своеобразный симбиоз, а в отдельных случаях как паразитизм.
Чрезвычайно сложные отношения сформировались у муравьев с некоторыми грибами. Обитающие в Центральной и Южной Америке муравьи-листорезы (Atta, Acromyjmex) срезают челюстями кусочки листьев, но не питаются ими и не строят из них жилье, а доставляют их в муравейник, тщательно пережевывают, смешивают с экскрементами и слюной и складывают в специальные камеры. На таких своеобразных грядках вырастают гифы грибов. На свободных концах гиф образуются многочисленные скопления коротких и вздутых ветвей, называемых кольраби. Они и служат пищей муравьям. Молодые самки, покидая муравейник, забирают с собой кусочки грибницы и при постройке нового муравейника сразу же приступают к возделыванию «грибных садов».
Известно до 100 видов таких муравьев. Очень мелкие Cyphomyrmex rhimosus поселяются в пустых раковинах или во вздутых листьях растений, а грибную плантацию размещают глубоко под землей, где выращивают желтые дрожжи, удобряя грядки экскрементами гусениц. Другие виды цифомирмексов возделывают грибные сады на компосте из древесной пыли и экскрементов жуков-дровосеков. Экскрементами гусениц удобряют свои плантации также муравьи Trachimyrmex и Apterostigma.
Установлено, что в гнездах муравьев произрастают такие грибы, как Hypomyces и шляпочный гриб Rhozites gongylophora, которые больше нигде не обнаружены. Только в муравейниках некоторых муравьев рода Lasius найден гриб Septosporium; гриб Leicosporium известен лишь из гнезд муравья Cyphomyrraex costatus. Все это позволяет сделать предположение, что в грибных садах муравейников в процессе естественного отбора образовались особые виды «муравьиных грибов».
Своеобразные грибные плантации встречаются также в гнездах термитов, где на специально откладываемых скоплениях экскрементов и кусочков древесины разводятся различные плесневые грибы. Но иногда, как и у муравьев, в них встречаются специфические грибы, которые больше нигде не произрастают (например, Termito-myces).
Грибные сады встречаются и у некоторых жесткокрылых. Так, самка непарного короеда (Xyleborus dispar), покидая гнездо, в специальных «карманчиках» уносит кусочки грибницы. Прогрызая ходы в новом дереве, короед образует там грибные камеры и рассеивает в них грибницу. Жук поддерживает в этих камерах определенную влажность – смачивает грибные грядки особыми выделениями, которые стимулируют рост полезных грибов и уничтожают сорняки. Личинки короеда питаются не древесиной, а структурами белого цвета, созревающими на свободных концах гиф гриба и напоминающими «муравьиные кольраби». Эти грибы нигде, кроме жилищ короедов, не растут. Однако, если из гнезда удалить самку, ухаживающую за грибным садом, то вскоре возделываемые грибы зарастают сорными грибами и погибают. Вслед за ними погибают от голода и личинки короеда. Такой своеобразный симбиоз обнаружен пока у небольшого количества видов из родов Anisandrus, Xyleborus, Xylotrupes, а также у некоторых ложнокороедов (Bostrychidae) и сверлил (Lymexylonidae).
Известны также комары родов Asphondylia, Lasioptera, Schizomia, Dasyneura, Contarinia, культивирующие грибы в галлах на зонтичных и бобовых растениях.
Между насекомыми и грибами также складываются взаимоотношения типа симбиоза, т. е. взаимовыгодного сожительства. Грибы дают насекомым богатую белками пищу, а взамен получают все необходимые для роста и развития условия. Здесь, как и в муравейниках или термитниках, налицо тесная взаимозависимость, невозможность раздельного существования ни насекомых, ни растений.
Роль травоядных животных в степных, луговых и тундровых биоценозах. Общий облик растительного покрова зависит не только от почвенно-климатических условий, но и от деятельности животных, находящихся в определенной связи с растениями. Изменение животного населения в любом из ландшафтов приводит к перемене в растительности. Полное же исключение животных сопровождается гибелью сообщества. Как отмечает Н. П. Наумов, в понятия «степь», «тундра», «лес», «луг» необходимо включать и животное население, и растительность потому, что они не только живут совместно, но и нуждаются друг в друге.
Широко известна роль слепушонков (Ellobius) в глинистых пустынях-такырах. Роясь в почве, они выбрасывают на поверхность холмики разрыхленной земли. На этих холмиках прорастают семена растений, которые в обычных условиях такыров не в состоянии пробиться через твердый слой засохшей глины. Постепенно растения покрывают некогда безжизненные пространства, создавая благоприятные условия для других растений и животных. Следовательно, слепушонки изменяют ландшафт, способствуют озеленению такыров.
Отмечаются случаи, когда истребление копытных в степях приводило к перерождению там растительности. Оказывается, многие злаки, основные степные растения, успешно развиваются и растут лишь при условии, если их объедают, «подстригают», копытные. Как только это исключается, они начинают вырождаться и в растительном сообществе происходит глубокая перестройка. Становится очевидным, что между копытными и злаками исторически сложились определенные межвидовые взаимоотношения. Благодаря им растения и животные приспособились к совместному существованию. На основе этого сосуществования и взаимного влияния и сформировался характерный степной биоценоз. Естественно, рассматривать копытных и злаки изолированно, вне связи с другими видами растений и животных, можно только условно, но доминирование взаимоотношений этих компонентов степного биогеоценоза не вызывает сомнения.
Примерно такую же роль играют копытные на полупустынных и пустынных пастбищах. Если они периодически не выедают полыней, то последние стареют и отмирают, а типичный ландшафт с определенным растительным покровом быстро перерождается. Такое явление характерно и по отношению к другим растительным ассоциациям. Повреждая растения неравномерно, избирательно, животные поддерживают тем самым нормальное соотношение их в сообществе.
В любом биогеоценозе между количеством и качеством животного населения и растительности должна быть определенная пропорция. Если увеличивается количество фитофагов, они уничтожают больше растений и в конце концов остаются без пищи, обрекая самих себя на гибель. С. С. Шварц, базируясь на экспериментальных данных, указывает, что «...максимальная продукция северных травостоев отмечается не тогда, когда она полностью ограждена от потребителей, а тогда, когда «пресс консументов» достигает определенной нормы. Лишь когда численность грызунов приближается к 1000 на гектар, продукция травостоя опускается ниже «спокойной нормы».
Следовательно, травоядные животные являются не разрушителями естественных фитоценозов, а их созидателями, неотъемлемой составной частью любого ландшафта. В природе исторически выработались и функционируют механизмы, поддерживающие наиболее выгодные количественные соотношения численности травоядных животных и растений, служащих им пищей.