

Суммарное уравнение биосинтеза пальмитиновой кислоты.
1 ацетил-КоА+7малонил-КоА+14НАДФН2 → пальмитиновая к-та+8НS-КоА+14НАДФ+6Н2О+7СО2
Гидролиз ферментативного комплекса с синтезируемой ЖК происходит под действием пальмитатдеацилазы.
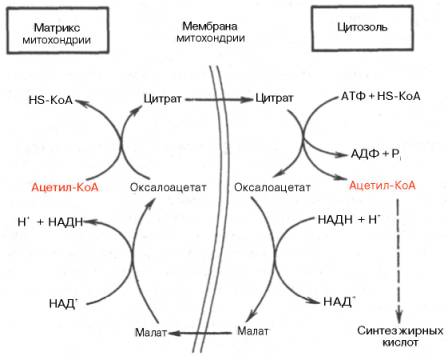
Удлинение ЖК происходит путем добавления ацетильных остатков в митохондриях и путем добавления малонильных остатков в цитоплазме.
ЛЕКЦИЯ №14
ТЕМА: ЛИПИДЫ III
Цель: Дать представление о механизмах регуляции липидного обмена, патологии, возникающие в результате его нарушения.
ПЛАН ЛЕКЦИИ
Регуляция обмена липидов. Регуляция окисления и синтеза ЖК.
Метаболизм кетоновых тел в норме и патологии
Транспортные липопротеины (ЛП)
Липопротеинемии и атеросклероз.
Депонирование и мобилизация жиров.
Распространение и функции Хс.
Синтез Хс.
Транспорт Хс.
Сложные липиды и миелинизация.
Регуляция обмена липидов
Интенсивность обмена липидов в тканях зависит от:
а) поступления липидов с пищей;
б) нервно-гумморальной регуляции
Избыточное поступление углеводов и ТАГ с пищей препятствует расходу эндогенных запасов ТАГ в жировой ткани. Прием даже полностью углеводной пищи существенно влияет на синтез ТАГ и Хс. Вместе с тем, растительные масла, содержащие ненасыщенные ЖК напротив, способствуют мобилизации липидов. Наличие липотропных факторов в пище облегчает биосинтез фосфолипидов, а их отсутствие способствует образованию ТАГ. Мобилизации ТАГ из жирового депо и угнетению биосинтеза холестерина способствуют голодание и эмоциональный стресс.
Нервно-гормональная регуляция липидного обмена
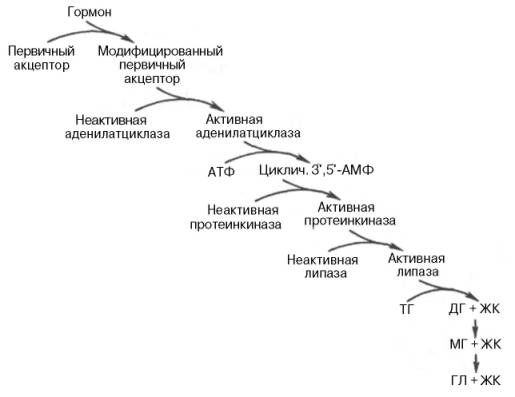
Стимуляторы тканевой липазы - адреналин, норадреналин, глюкагон, тироксин, адренокортикотропный гормон (активируют аденилатциклазу)
Ингибитор липолиза – инсулин (угнетает аденилатциклазу). В результате воздействия гормонов нервной клетки рецептор модифицирует свою структуру. В такой форме данный рецептор способен активировать аденилатциклазу, что стимулирует образование ц-АМФ и АТФ. Далее ц-АМФ активирует протеинкиназу, а последняя, путем фосфорилирования неактивную тканевую липазу превращает в активную.
Липолитический каскад (по Стайнбергу).
Окисление и синтез ЖК регулируется:
а) разделением этих противоположных процессов в пространстве (окисление ЖК в митохондриях, синтез ЖК в цитозоле)
б) резделением этих процессов во времени потеем аллостерической активации и ингибирования ферментов.
Метаболизм кетоновых тел в норме и патологии
Кетоновыми или ацетоновыми телами называются: ацетоацетат
β-гидроксибутират
ацетон
Кетогенез происходит в митохондриях печени. Наиболее значим гидроксиметил-глутаматный цикл.
Конденсация 2-х молекул ацетил-КоА с образованием ацетоацетил-КоА.
Е ацетил-КоА-ацетилтрансферазой
-
О
О
СН3─
C=О
║
║
│
СН3─
С ~ КоА +
СН3─
С ~ КоА→
СН2 +
НSКоА
│
О=
С ~ SКоА
Конденсация ацетоацетил-КоА еще с одной молекулой ацетил-КоА. Е гидроксиметилглутарил-КоА-синтетазы.
-
СН3─
C=О
О
СООН─
CН2
│
║
│
СН2 +
СН3─
С ~ КоА→
СН3─
С─ОН +
НSКоА
│
│
О=
С ~ SКоА
О=
С ~ SКоА
ацетоацетил-КоА
β-окси-β-метилглутарил-КоА (ГМГ)
Образовавшийся ГМГ под действием Е гидроксиметилглутарил-КоА-лиазы расщепляется на ацетоацетат и ацетил-КоА.
-
СООН─
CН2
СН3─
C=О
О
│
│
║
СН3─
С─ОН →
СН2 +
СН3─
С ~ КоА
│
│
О=
С ~ SКоА
О=
С─ОН
ГМГ
ацетоацетат
Ацетил-КоА вновь используется на первой стадии, т.е. замыкает процесс в цикле. Остальные кетоновые тела образуются из ацетоацетата:
β-гидроксибутират путем восстановления, Е НАД-зависимая гидроксибутират ДГ
ацетон в результате декарбоксилирования ацетоацетата, Е ацетоацетатдекарбоксилаза.
-
СН3─
C=О
О
│
→
║ + СО2
СН2
СН3─
С─ СН3 ацетон
│
О=
С─ОН +
НАДН2
→
НООС─

СН2─СН─ СН3
ацетоацетат
н ОН β-гидроксибутират
Для синтеза кетоновых тел используют ацетил-КоА, образовавшийся с помощью специфических регуляторных механизмов из ЖК! В печени кетоновые тела далее не превращаются, а поступают в кровь.
В норме кетоновые тела или отсутствуют или их концентрация невелика ≈ 3мг/дл (декалитр=10л).
Другие органы и ткани: сердце, легкие, почки, мышцы, нервная ткань – преобразуют кетоновые тела в ацетил-КоА, который сгорает в ЦТК до СО2 и Н2О, т.е. этими органами кетотела используются как источник энергии.
Содержание кетоновых тел в организме увеличивается при:
длительной мышечной работе,
голодании,
некоторых заболеваниях, например сахарный диабет.
Через двое суток голодания концентрация кетоновых тел в крови достигает 5-6мг/дл, через неделю 40-50мг/дл.
При сахарном диабете уровень кетоновых тел может достигать 300-400мг/дл, что приводит к метаболическому ацидозу, рН крови изменяется до 7,4 (±0,04), нарушаются функции мозга, вплоть до потери сознания.