

1 Стадия
1.дегидрирование
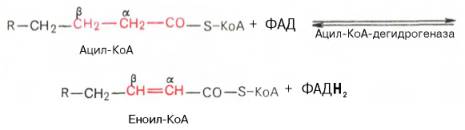
2. гидратация.
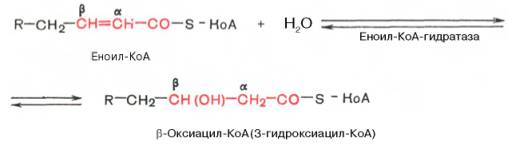
2 Стадия
3. дегидрирование
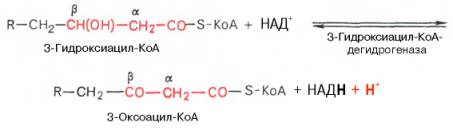 β-кетоацил
КоА
β-кетоацил
КоА
4.тиолазная реакция
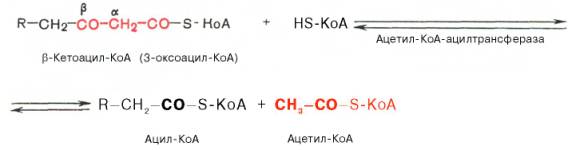
Происходит расщепление β-кетоацил КоА на ацил-КоА и ацетил-КоА, укоротившийся на два углеродных атома.
Далее цикл повторяется до тех пор, пока не образуется бутирил-КоА (4-углеродное соединение), который в свою очередь окисляется до 2 молекул ацетил-КоА.
Ацетил-КоА → в ЦТК
НАДН2,ФАДН2 → в дыхательную цепь
Энергетический баланс β-окисления (четное число атомов углерода)
При окислении жирной кислоты, содержащей n углеродных атомов, происходит n/2–1 цикл β-окисления (т.е. на один цикл меньше, чем n/2, так как при окислении бутирил-КоА происходит образование 2 молекул ацетил-КоА) и получится n/2 молекул ацетил-КоА. Следовательно, суммарное уравнение β-окисления пальмитиновой кислоты можно записать так:
Пальмитоил-КоА + 7ФАД + 7НАД+ + 7Н2O + 7HS-KoA ––> 8Ацетил-КоА + 7ФАДН2 + 7НАДН2.
При каждом цикле β-окисления образуются одна молекула ФАДН2 и одна молекула НАДН. Последние в процессе окисления в дыхательной цепи и сопряженного с ним фосфорилирования дают:
 7ФАДН2
х 2 = 14
7ФАДН2
х 2 = 14
7НАДН2 х 3 = 21 35 молекул АТФ ( в дыхательную цепь)
8 ацетил-КоА х 12 = 96 молекул АТФ (в ЦТК)
ИТОГО: 35+96=131-1(на образование активной формы жк) = 130 молекул АТФ.
Для ненасыщенной жк кол-во двойных связей х 2 = количество молекул АТФ.
Изменение свободной энергии ΔF при полном сгорании 1 моля пальмитиновой кислоты составляет 2338 ккал, а богатая энергией фосфатная связь АТФ характеризуется величиной 7,6 ккал/моль. Примерно 990 ккал (7,6 х 130), или 42% от всей потенциальной энергии пальмитиновой кислоты при ее окислении в организме, используется для ресинтеза АТФ, а оставшаяся часть, теряется в виде тепла. |
Окисление жирных кислот с нечетным количеством атомов углерода.
Жирные кислоты с нечетным числом углеродных атомов окисляются таким же образом, как и жирные кислоты с четным числом углеродных атомов, с той лишь разницей, что на последнем этапе расщепления (β-окисления) образуется одна молекула пропионил-КоА и одна молекула ацетил-КоА, а не 2 молекулы ацетил-КоА.
Активированный пропионил-КоА – включается в цикл трикарбоновых кислот после превращения в сукцинил-КоА.
1.карбоксилирование
2. изомеризация
ЦТК |
|
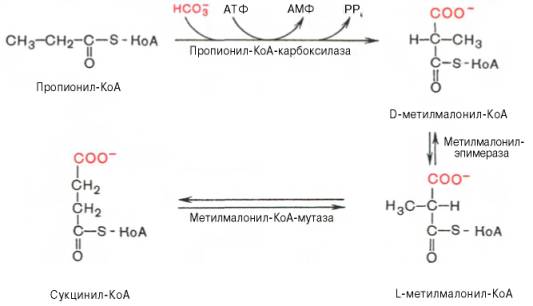
При недостатке витамина В12 последняя реакция замедляется и с мочой выводится большое количество метилмалоната и пропионата.
Особенности окисления ненасыщенных жиных кислот
До двойных связей окисление ненасыщенных жирных кислот происходит так же, как окисление насыщенных жк.
|
Н |
О |
|
Если двойные связи имеют ту же конформацию, что в еноил-КоА (транс),то окисление идет обычным путем. Если же конфигурация ненасыщенных ЖК «цис», то в реакции действует дополнительный фермент ∆ 3,4-цис-∆ 2,3-трансеноилизомераза, который перемещает двойную связь из цис в транс положение. Скорость окисления ненасышенных жирных кислот выше, чем у насышенных. |
3 |
│2 |
║1 |
|
|
─ С ═ |
С ─ |
С ~ |
S ─ КоА |
|
│ |
|
|
(∆ 2,3) |
|
Н |
|
|
||
Н |
Н |
|
О |
|
│4 |
│3 |
2 |
║1 |
|
─ С ═ |
С ─ |
СН2 |
─ С ~ |
S ─ КоА (∆ 3,4) |