

Лекция 5.
Общий путь катаболизма как источник субстратов тканевого дыхания – окислительное декарбоксилирование пирувата, цикл трикарбоновых кислот. Подведение итога: образование АТФ методом субстратного и окислительного фосфорилирования АДФ
Общий путь катаболизма включает: 1) окислительное декарбоксилирование пирувата до ацетил-КоА; 2) окисление ацетильной группы ацетил-КоА (2 и 3 углеродные атомы пирувата) в цикле трикарбоновых кислот; 3) выделение и аккумулирование энергии при дегидрировании метаболитов общего пути катаболизма в митохондриальных цепях переноса электронов.
Пируват образуется из углеводов (глюкоза), глицерола, гликогенных аминокислот и лактата. Ацетил-КоА занимает центральное место в общем пути катаболизма и образуется в митохондриях: 1) при окислительном декарбоксилировании пирувата; 2) при β-окислении жирных кислот; 3) из кетогенных аминокислот.
1. Окислительное декарбоксилирование пировиноградной кислоты
Пировиноградная кислота связывает гликолиз с циклом трикарбоновых кислот. Пируват переносится из цитозоля в матрикс митохондрий с помощью переносчика по механизму симпорта с протоном. В матриксе митохондрий пируват превращается в ацетил-КоА. Этот процесс называется окислительное декарбоксилирование пирувата и катализируется пируватдегидрогеназным комплексом (пируватдегидрогеназной системой). Высокая концентрация пируватдегидрогеназного комплекса обнаружена в сердечной мышце и почках.
Пируватдегидрогеназный комплекс является классическим мультиферментным комплексом, в котором промежуточные продукты остаются связанными на поверхности молекулы фермента до образования конечного продукта.
В состав пируватдегидрогеназного комплекса входит 3 фермента и 5 кофакторов. I-й фермент – пируватдегидрогеназа содержит кофактор тиаминпирофосфат (производное витамина В1); II-й фермент – дигидролипоилтрансацетилаза содержит кофакторы липоевую кислоту (ЛК, 6,8-дитиооктановая кислота) и кофермент А (НS-КоА), причем остаток липоевой кислоты присоединен к апоферменту путем образования амидной связи между карбоксильной группой ЛК и ε-аминогруппой лизина белка (образуется длинная «рука», состоящая из 13 атомов углерода); III-й фермент - дигидролипоилдегидрогеназа содержит кофакторы ФАД и НАД+.

2. Механизм окислительного декарбоксилирования пирувата
I этап. Пируват взаимодействует с кофактором пируватдегидрогеназы тиаминпирофосфатом. Основную роль играет второй углеродный атом тиазольного кольца ТПФ, который легко теряет протон, превращаясь в карбанион. Карбанион атакует частично положительно заряженный α-углеродный атом пирувата с возникновением связи С−С. Сильно электрофильный атом азота в карбоксиэтил-ТПФ способствует его декарбоксилированию с образованием гидроксиэтил-ТПФ.

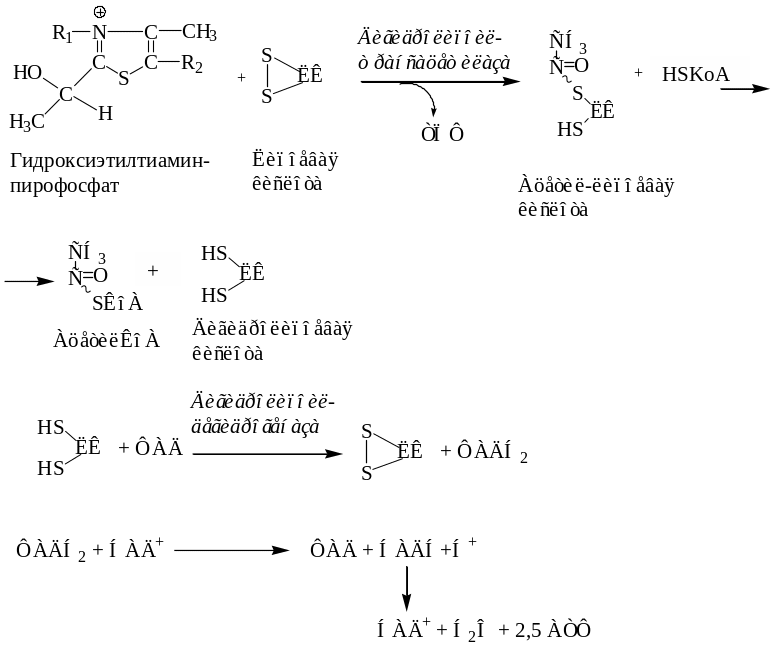
II этап. В следующей реакции, катализируемой дигидролипоилтрансацетилазой, гидроксиэтил-ТПФ взаимодействует с липоевой кислотой. Происходит перенос гидроксиэтильной группы на один из атомов серы ЛК. При этом гидроксиэтильная группа окисляется в ацетильную. В процессе окисления гидроксиэтильной группы и восстановления SH-группы ЛК возникает макроэргическая связь. Затем ацетильный остаток переносится на второй кофермент дигидролипоилтрансацетилазы – HS-KoA, а ЛК полностью восстанавливается. Образованный ацетил-КоА отделяется от полиферментного комплекса.
III этап. Восстановленная форма ЛК окисляется дигидролипоилдегидрогеназой.
3. Регуляция пируватдегидрогеназного комплекса. Превращение пирувата в ацетил-КоА – процесс необратимый. Поэтому синтез глюкозы из ацетил-КоА невозможен. Обычно ацетил-КоА далее превращается 2-мя путями: 1) ацетильная группа ацетил-КоА окисляется до СО2 и Н2О через ЦТК и сопряженные цепи переноса электронов с выделением и запасанием энергии в виде АТФ; 2) используется для синтеза кетоновых тел, холестерола и жирных кислот.
Пируватдегидрогеназный комплекс регулируется методом фосфорилирования-дефосфорилирования. Повышение величин отношений НАДН/НАД+, ацетил-КоА/КоА или АТФ/АДФ способствует фосфорилированию фермента протеинкиназой и дезактивации комплекса. Следовательно, пируватдегидрогеназный комплекс инактивируется, если клетка богата энергией и биосинтетическими предшественниками.
Пируват и АДФ, наоборот, активируют пируватдегидрогеназный комплекс посредством ингибирования протеинкиназы.
Вазопрессин активирует пируватдегидрогеназный комплекс путем повышения концентрации ионов кальция в митохондриях, которые активируют протеинфосфатазу (дефосфорилирование фермента). Инсулин также усиливает превращение пирувата в ацетил-КоА через дефосфорилирование пируватдегидрогеназного комплекса.