

5. Глюконеогенез
Глюконеогенез – синтез углеводов из неуглеводных предшественников (гликогенных аминокислот, глицерола, лактата, пропионовой кислоты). Глюконеогенез протекает в цитозоле клеток печени и почек, хотя некоторые предшественники образуются в митохондриях. Глюконеогенез необходим для синтеза глюкозы, которая является источником энергии, особенно для нервной ткани и эритроцитов. При голодании более 1 суток за счет глюконеогенеза поддерживается базальный уровень глюкозы в крови, и метаболизм обеспечивается промежуточными продуктами ЦТК. При интенсивной физической работе синтез глюкозы происходит из лактата и глицерола; при голодании – из глицерола, образующегося при распаде жиров и аминокислот, освобождающихся в процессе катаболизма белков мышц.
Большинство стадий глюконеогенеза представляет собой обращение реакций гликолиза. Величина G0 при образовании пирувата из глюкозы в стандартных условиях клетки равно -20 ккал/моль (-84 кДж/моль). Основное уменьшение свободной энергии имеет место в трех необратимых реакциях, катализируемых гексокиназой, фосфофруктокиназой и пируваткиназной:
В процессе глюконеогенеза для обращения этих трех необратимых этапов используются другие ферменты.
1. При интенсивной мышечной работе образуется лактат, который током крови поступает в печень. В цитозоле под действием лактатдегидрогеназы лактат окисляется в пируват, который поступает в митохондрии.

Прямой синтез фосфоенолпирувата из пирувата невозможен из-за необратимости пируваткиназной реакции, поэтому используется обходной путь из нескольких ферментов. Вначале пируват поступает в митохондрии и при участии биотин-зависимой пируваткарбоксилазы, СО2 и АТФ карбоксилируется с образованием оксалоацетата. Это ключевой момент для энергетического преодоления пируваткиназной реакции в обходных реакциях.
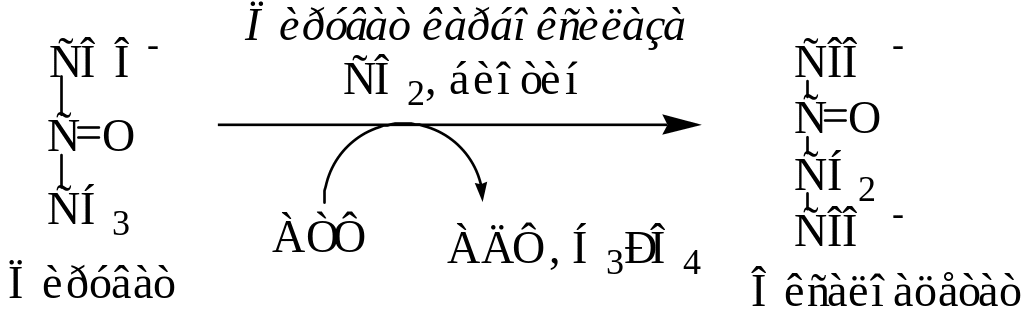
Для транспорта из митохондрий в цитозоль оксалоацетат восстанавливается в малат с участием фермента НАД-зависимой малатдегидрогеназы.

В цитозоле происходит обратный процесс окисления малата в оксалоацетат под действием цитозольной малатдегидрогеназы с коферментом НАД+.

Затем оксалоацетат в результате декарбоксилирования и фосфорилирования под влиянием фермента фосфоенолпируваткарбоксикиназы превращается в фосфоенолпируват. Донором фосфатного остатка служит ГТФ. На этом завершается путь обхода необратимой пируваткиназной реакции.

Фосфоенолпируват в результате ряда обратимых реакций гликолиза превращается во фруктозо-1,6-бисфосфат.
2.. Превращение фруктозо-1,6-бисфосфата во фруктозо-6-фосфат катализируется специфической фосфатазой – фруктозо-1,6-бисфосфатазой (обход необратимой фосфофруктокиназной реакции).
3. В последующей обратимой стадии биосинтеза глюкозы фруктозо-6-фосфат превращается в глюкозо-6-фосфат, который дефосфорилируется под влиянием фермента глюкозо-6-фосфатазы (обход необратимой гексокиназной реакции).
В большинстве тканей на этом завершается глюконеогенез и свободная глюкоза не выделяется. Глюкозо-6-фосфат далее превращается разными путями, в том числе и в гликоген. Только два органа – печень и почки, в которых имеется фермент глюкозо-6-фосфатаза, связанный с мембранами эндоплазматического ретикулума, способны освобождать глюкозу в кровь.
В организме существует взаимосвязь гликолиза и глюконеогенеза на органном уровне. Основным субстратом для глюконеогенза является лактат, образованный активной скелетной мышцей. Плазматическая мембрана обладает высокой проницаемостью для лактата. Поступив в кровь, лактат переносится в печень, где в цитозоле окисляется в пируват. Пируват затем превращается в глюкозу по пути глюконеогенеза. Глюкоза поступает далее в кровь и поглощается скелетными мышцами. Эти превращения составляют цикл Кори.