

3.6. Энергетический баланс аэробного распада глюкозы
1. В специфическом пути распада глюкозы образуется 2 молекулы пирувата, 2 АТФ (субстратное фосфорилирование) и 2 молекулы НАДН+Н+.
2. Окислительное декарбоксилирование каждой молекулы пирувата - 2,5 АТФ; декарбоксилирование 2 молекул пирувата дает 5 молекул АТФ.
3. В результате окисления ацетильной группы ацетил-КоА в ЦТК и сопряженных ЦПЭ – 10 АТФ; 2 молекулы ацетил-КоА образуют 20 АТФ.
4. Малатный челночный механизм переносит НАДН+Н+ в митохондрии – 2,5 АТФ; 2 НАДН+Н+ образуют 5 АТФ.
Итого: при распаде 1 молекулы глюкозы в аэробных условиях образуется 32 молекулы АТФ.
В тканях, где действует глицерофосфатный челночный механизм переноса восстановительных эквивалентов с помощью глицерофосфатдегидрогеназы, кофактором которой является ФАД, в митохондрии образуется 2 молекулы ФАДН2, которые в ЦПЭ дадут 3 молекулы АТФ. Тогда при полном окислении глюкозы будет образовываться 30 АТФ.
Аэробный распад 1 молекулы глюкозы до СО2 и Н2О сопряжен с аккумулированием 30-32 молекул АТФ
3.7. Метаболизм глюкозы в анаэробных условиях
В анаэробных условиях распад глюкозы до пирувата происходит аналогично, как и в аэробных условиях.
Судьба пирувата, образуемого при гликолизе, зависит от наличия или отсутствия кислорода. В анаэробных условиях в результате одиннадцатой реакции происходит восстановление пировиноградной кислоты в молочную кислоту (лактат). Реакция протекает при участии фермента лактатдегидрогеназы и кофермента НАДН+Н+, образованного при окислении глицеральдегид-3-фосфата в 1,3-бисфосфоглицериновую кислоту. Образование лактата позволяет регенерировать НАД+, который затем снова используется в реакции, катализируемой глицеральдегид-3-фосфатдегидрогеназой.
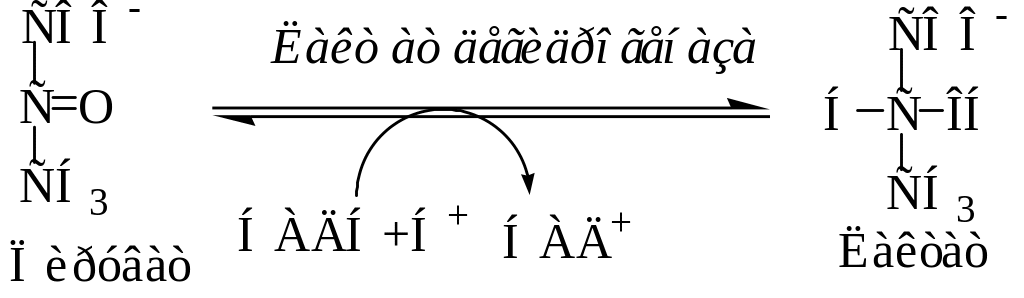
В гликолизе выделяют центральную окислительно-восстановительную реакцию гликолиза (гликолитическую оксидоредукцию), которая состоит в окислении глицеральдегид-3-фосфата с образованием НАДН+Н+ и последующим переносом атомов водорода от НАДН+Н+ на пируват.
Значение гликолиза велико в тканях и органах, где ограничен доступ кислорода или возможно внезапное и резкое возрастание скорости потребления АТФ. Например, в работающей сердечной мышце скорость потребления АТФ может возрасти в несколько раз, а в работающей скелетной мышце – в несколько десятков раз. Когда аэробных возможностей для обеспечения повышенной потребности в АТФ становится недостаточно, дополнительное количество АТФ будет черпаться из гликолиза.
Гликолиз до лактата происходит только в анаэробных условиях и в клетках, лишенных митохондрий (эритроциты).
Энергетическая эффективность гликолиза в анаэробных условиях составляет 2 молекулы АТФ на одну молекулу глюкозы
3.7.1. Гликогенолиз. Анаэробному расщеплению могут подвергаться глюкозные остатки, отщепленные от концов цепей гликогена фосфоролитическим путем – гликогенолиз. Включение гликогена в процесс анаэробного распада осуществляется с помощью трех ферментов: 1) гликоген-фосфорилаза катализирует расщепление концевых α-1,4-гликозидных связей с выделением глюкозо-1-фосфата; 2) дебраншинг фермент катализирует расщепление α-1,6-гликозидных связей с выделением глюкозо-1-фосфата; 3) фосфоглюкомутаза катализирует превращение глюкозо-1-фосфата в глюкозо-6-фосфат. Поскольку при гликогенолизе на образование глюкозо-6-фосфата АТФ не тратится, в процессе гликогенолиза накапливаются не две, а три молекулы АТФ на каждый отделенный глюкозный остаток. Однако это не означает, что гликогенолиз энергетически более выгоден, чем гликолиз, так как на синтез гликогена из глюкозы также расходуется АТФ.
3.7.2. Эффект Пастера - ингибирование гликолиза (накопления лактата) кислородом. Механизмы эффекта:
1. При аэробном распаде углеводов не функционирует окислительно-восстановительная реакция гликолиза, поскольку восстановительные эквиваленты НАДН+Н+ из цитоплазмы переносятся в митохондрии. Поэтому НАДН+Н+ не может служить для восстановления пирувата в лактат.
2. При наличии аэробных условий образуются цитрат и АТФ, которые аллостерически тормозят активность фосфофруктокиназы (реакция лимитирует скорость всего процесса).
3. Дыхание подавляет гликолиз из-за конкуренции за АДФ между митохондриальной цепью переноса электронов, с одной стороны, и фосфоглицераткиназной и пируваткиназной реакциями гликолиза в цитоплазме, с другой. АДФ преимущественно используется в реакции окислительного фосфорилирования в митохондриях, а не субстратного фосфорилирования в гликолизе.
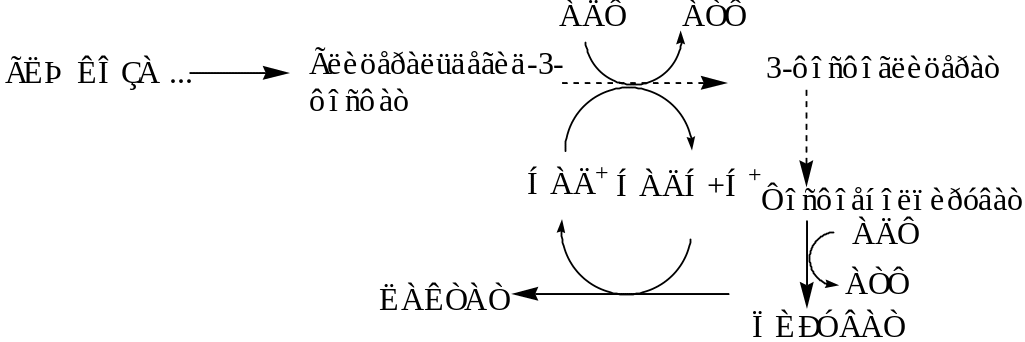
3.7.3. Спиртовое брожение. Спиртовое брожение осуществляется так называемыми дрожжеподобными организмами, а также некоторыми плесневыми грибами. Суммарную реакцию спиртового брожения можно изобразить следующим образом:
С6Н12О6 → 2 С2Н5ОН + 2 СО2
Процесс спиртового брожения протекает по пути гликолиза до образования пирувата.
В дрожжевых клетках (спиртовое брожение) пируват вначале подвергается декарбоксилированию, в результате чего образуется ацетальдегид. Данная реакция катализируется пируватдекарбоксилазой, которая требует наличия ионов Mg2+ и кофермента тиаминпирофосфата:
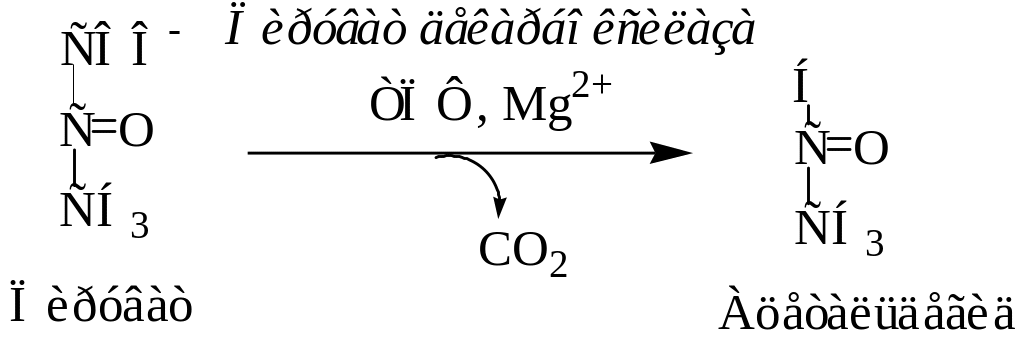
Затем образовавшийся ацетальдегид присоединяет к себе водород, отщепляемый от НАДН+Н+, восстанавливаясь при этом в этанол. Реакция катализируется ферментом алкогольдегидрогеназой.

Таким образом, конечными продуктами спиртового брожения являются этанол и СО2, а не молочная кислота, как при гликолизе.
4. Пентозофосфатный путь распада глюкозы (ПФП) называется также гексозомонофосфатным шунтом или фосфоглюконатным путем. Этот альтернативный гликолизу и ЦТК путь окисления глюкозы был описан в 50-х годах ХХ века Ф.Дикенсом, Б.Хорекером, Ф.Липманном и Е.Рэкером.
Ферменты пентозофосфатного пути локализованы в цитозоле. Наиболее активно ПФП протекает в почках, печени, жировой ткани, коре надпочечников, эритроцитах, лактирующей молочной железе. В большинстве из этих тканей протекает процесс биосинтеза жирных кислот и стероидов, что требует НАДФН.
Выделяют две фазы ПФП: окислительную и неокислительную (рис. 3).
ADPH
and Synthesizes Five-Carbon Sugars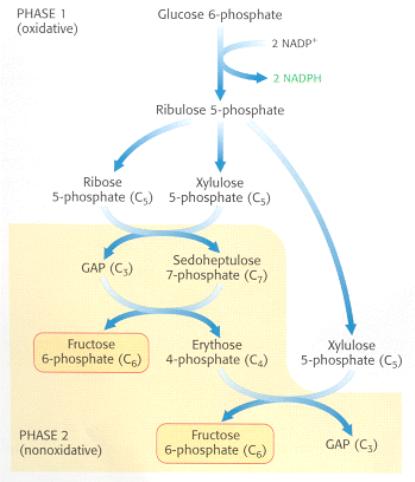
Рис. 5. Пентозофосфатный путь обмена углеводов. Состоит из окислительной фазы образования НАДФН и неокислительной фазы превращений фосфорилированных
Подписи сверху вниз: Фаза 1 (окислительная); глюкозо-6-фосфат; 2 НАДФ+; 2 НАДФН; Рибулозо-5-фосфат; Рибозо-5-фосфат (С5); Ксилулозо-5-фосфат (С5); 3-ФГА (С3); седогептулезо-7-фосфат (С7); Фруктозо-6-фосфат (С6); Эритрозо-4-фосфат (С4); ксилулозо-5-фосфат (С5); Фаза 2 (неокислительная); Фруктозо-6-фосфат (С6); 3-ФГА (С3).