

24.Тетанус (тетаническое мышечное сокращение)
При слиянии одиночных мышечных сокращений во время ритмического раздражения наблюдается тетанус (тетаническое сокращение). При низких частотах раздражения механический ответ может быть волнообразным, так как волокно частично расслабляется в промежутках между стимулами, - это зубчатый тетанус. Если частоту раздражения повысить, получается гладкий тетанус без осцилляции
25.Механизмы сокращения мышечного волокна
В покоящихся мышечных волокнах при отсутствии импульсации мотонейрона поперечные миозиновые мостики не прикреплены к актиновым миофиламентам. Тропомиозин расположен таким образом, что блокирует участки актина, способные взаимодействовать с поперечными мостиками миозина. Тропонин тормозит миозин — АТФ-азную активность и поэтому АТФ не расщепляется. Мышечные волокна находятся в расслабленном состоянии.
При сокращении мышцы длина А-дисков не меняется, J-диски укорачиваются, а Н-зона А-дисков может исчезать (рис. 4.3.).
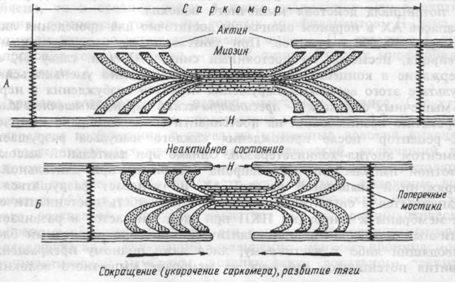
Рис.4.3. Сокращение мышцы. А — Поперечные мостики между актином и миозином разомкнуты. Мышца находится в расслабленном состоянии. Б — Замыкание поперечных мостиков между актином и миозином. Совершение головками мостиков гребковых движений по направлению к центру саркомера. Скольжение актиновых нитей вдоль миозиновых, укорочение саркомера, развитие тяги.
Эти данные явились основой для создания теории, объясняющей сокращение мышцы механизмом скольжения (теорией скольжения) тонких актиновых миофиламентов вдоль толстых миозиновых. В результате этого миозиновые миофиламенты втягиваются между окружающими их актиновыми. Это приводит к укорочению каждого саркомера, а значит, и всего мышечного волокна.
Молекулярный механизм сокращения мышечного волокна состоит в том, что возникающий в области концевой пластинки потенциал действия распространяется по системе поперечных трубочек вглубь волокна, вызывает деполяризацию мембран цистерн саркоплазмати-ческого ретикулума и освобождение из них ионов кальция. Свободные ионы кальция в межфибриллярном пространстве запускают процесс сокращения. Совокупность процессов, обуславливающих распространение потенциала действия вглубь мышечного волокна, выход ионов кальция их саркоплазматического ретикулума, взаимодействие сократительных белков и укорочение мышечного волокна называют «электромеханическим сопряжением».Временная последовательность между возникновением потенциала действия мышечного волокна, поступлением ионов кальция к миофибриллам и развитием сокращения волокна показана на рисунке 4.4.
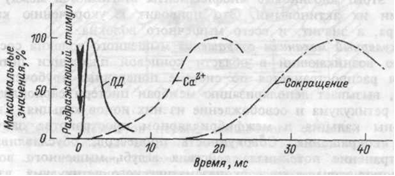
Рис.4.4. Схема временной последовательности развития потенциала действия (ПД), освобождения ионов кальция (Са2+) и развития изометрического сокращения мышцы.
При концентрации ионов Са2+ в межмиофибриллярном пространстве ниже 10″ тропомиозин располагается таким образом, что блокирует прикрепление поперечных миозиновых мостиков к нитям актина. Поперечные мостики миозина не взаимодействуют с нитями актина. Продвижение относительно друг друга нитей актина и миозина отсутствует. Поэтому мышечное волокно находится в расслабленном состоянии. При возбуждении волокна Са2+ выходит из цистерн саркоплазматического ретикулума и, следовательно, концентрация его вблизи миофибрилл возрастает. Под влиянием активирующих ионов Са2+ молекула тропонина изменяет свою форму таким образом, что выталкивает тропомиозин в желобок между двумя нитями актина, освобождая тем самым участки для прикрепления миозиновых поперечных мостиков к актину. В результате поперечные мостики прикрепляются к актиновым нитям. Поскольку головки миозина совершают «гребковые» движения в сторону центра саркомера происходит «втягивание» актиновых миофиламентов в промежутки между толстыми миозиновыми нитями и укорочение мышцы.
Электромеханическое сопряжение - это последовательность процессов, в результате которых потенциал действия плазматической мембраны мышечного волокна приводит к запуску цикла поперечных мостиков .
Сила сокращения изолированной скелетной мышцы при прочих равных условиях зависит от исходной длины мышцы. Умеренное растяжение мышцы приводит к тому, что развиваемая ею сила возрастает по сравнению с силой, развиваемой нерастянутой мышцей. Происходит суммирование пассивного напряжения, обусловленного наличием эластических компонентов мышцы, и активного сокращения. Максимальная сила сокращения достигается при размере саркомера 2—2,2 мкм (рис. 2.26). Увеличение длины саркомера приводит к уменьшению силы сокращения, поскольку уменьшается область взаимного перекрытия актиновых и миозиновых нитей. При длине саркомера 2,9 мкм мышца может развивать силу, равную только 50% от максимально возможной.