

1.2.5. Комплексины и их возможное участие в глутаматной эксайтотоксичности
Комплексины – это семейство специфических эволюционно консервативных белков цитоплазмы нейронов, связывающихся с рецептором белка а-SNAP – белковым комплексом SNARE, состоящим из синтаксина, синаптобревина и SNAP-25 и обеспечивающим высвобождение медиаторов из синаптических пузырьков в синаптическую щель. Известно, что существует два типа комплексинов – CPLX1 и CPLX2. Первый представлен в тормозящих (ингибирующих) синапсах, второй – в возбуждающих [119], [120]. Это сравнительно небольшие по размеру (молекулярная масса около 19000 Да) белки цитозоля нейронов. Комплексины богаты остатками глутаминовой кислоты и лизина [121]. В основном локализуются в пресинаптических окончаниях нейронов млекопитающих. Белковая молекула состоит из 4 функциональных частей:
- N-концевой домен,
- вспомогательная спираль,
- центральная спираль,
- С-концевой домен [122].
Комплексины выполняют двойную функцию: они могут действовать и как активаторы, и как ингибиторы экзоцитоза в зависимости от физиологического состояния клетки. Ингибирование слияния мембран необходимо для предотвращения самопроизвольного экзоцитоза в синапсе [123], [124]. Последние данные поддерживают версию, что синаптотагмин играет роль в конформационных изменениях SNARE при изменении концентрации кальция [123]. Связывание кальция с синаптотагмином провоцирует изменение конформации и вытеснение комплексина, после чего мембрана везикулы сливается с пресинаптической мембраной и происходит экзоцитоз [125]. При низкой концентрации кальция экзоцитоз-ингибирующий эффект комплексина выражен сильнее. Он снижается посредством синаптотагмина при растущей концентрации кальция с переводом синаптотагмина в активное состояние [123].
Исследования, проведённые Н.М. Раевской и др., обнаружили у человека два варианта транскриптов гена Cplx2, различающиеся 5’- концевыми участками (CPLX_v1 и CPLX_v2). Они имеют высокое сходство с аналогичными транскриптами у крыс (Cplx_v1 и Cplx_v2) [126]. Этот случай не уникальный, он известен для многих генов, причём различные транскрипты одного и того же гена часто являются тканеспецифичными либо появляются в одной и той же ткани и клетке в зависимости от стадии клеточного цикла.
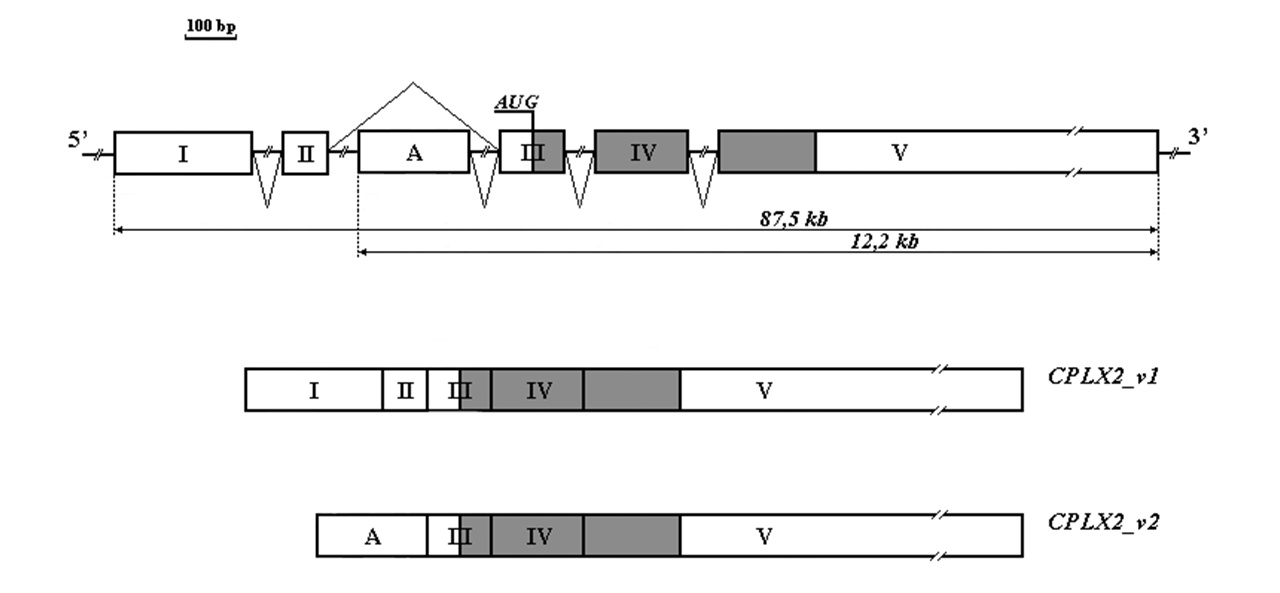
Рисунок 1.1. Структурная организация гена CPLX2 человека и его транскриптов
Факторы, определяющие транскрипцию по одному или по другому типу, до конца не выяснены. Вероятно, роль в «выборе» того или иного продукта играют альтернативные промоторы и взаимодействующие с ними факторы, что определяется потребностью клетки в том или ином продукте.
Исследования экспрессии гена Cplx2 также показали двустороннюю роль комплексинов в механизме экзоцитоза. Так, например, на культуре клеток РС12 было обнаружено, что Cplx2 стимулирует экзоцитоз при низком экспрессионном уровне и ингибирует при более высоком [127]. Изменение экспрессии гена этого белка при ишемии мало изучено, хотя известно, что важнейшие звено в механизме развертывания ишемического повреждения ткани мозга – глутаматная эксайтотоксичность – частично обеспечивается высвобождением глутамата в составе синаптических везикул. Также показано, что Cplx2 активно экспрессируется не только в нейронах, но и в астроцитах, которые принимают важное участие в регуляции оборота глутамата [128]. Таким образом, белок CPLX2 может принимать важное участие в глутаматной эксайтотоксичности. Например, было показано, что увеличение экспрессии Cplx2 приводит к нарушениям высвобождения глутамата с последующими патологическими изменениями [129], [130]. Также следует отметить, что функции комплексинов связаны с регуляцией экзоцитоза не только глутамата, но и других важных медиаторов и трофических факторов.